

Automation of Auditory Cognitive Processing in 100 m Sprint Elites' Expertise Domain: An ERP Study
-
摘要:目的
基于100 m跑起跑听觉专项认知加工任务,探讨精英运动员专长领域听觉认知加工自动化的事件相关电位(Event-Related Potential,ERP)特征,分析精英运动员和普通人大脑皮层活动的差异。
方法采用ERP技术和感知-认知范式,采集被试对100 m起跑三阶段听觉信号探测的ERP特征。
结果①在“各就位”和“预备”阶段,精英运动员组的P2、P3、N1振幅显著低于普通组;②在起跑鸣枪阶段,精英运动员组的P2振幅显著高于普通组;③精英运动员组在次要刺激阶段(“各就位”和“预备”阶段),额顶网络活动显著减少,在起跑鸣枪阶段,额顶网络活动显著增强。
结论①精英运动员的听觉认知加工自动化体现出无意识特征,加工过程很少或根本不消耗意识资源,普通人的意识加工更多;②支持当前认知加工自动化理论中的渐进观,即不存在一个绝对自动化的阶段,受控加工也参与自动化加工的启动阶段,精英运动员在处理认知信号时会基于认知任务差异,在受控加工和自动化加工之间进行转换;③精英运动员认知加工自动化还呈现出半球专属特有启动(hemisphere-specific priming)特征,左半球涉及意识的言语控制活动受到抑制和减少,同时涉及动作技能表现的右半球活动显著增强。
Abstract:ObjectiveBased on the task of auditory cognitive processing for 100 m sprint elites, the characteristics of the event-related potential (ERP) of the auditory cognitive automatic processing of elite athletes were investigated, and the differences of cerebral cortex activity between elite athletes and novice athletes were analyzed.
MethodsThe ERP technology and perceptual-cognitive paradigm were used to collect the ERP characteristics of the three-stage auditory signal detection at the start of 100 m.
Results① The amplitudes of P2, P3 and N1 in the elite group were significantly lower than those in the novice group on the stage of "On Your Marks" and "Set" stage. ② On the "Go" stage, P2 amplitude of elite group was significantly higher than that of novice group. ③ In the elite group, fronto-parietal network activity decreased significantly on the secondary stimulus stage (each on the stage of "On Your Marks" and "Set"), but increased significantly on the "Go" stage.
Conclusions① The auditory cognitive automatic processing of elite athletes shows unconsciousness, consuming little or no conscious resources, while the novice athletes consume more resources. ② The results support the gradual view in the current automatic processing theory, that is, there is no absolute automatic stage, and controlled processing also participates on the start-up stage of automatic processing. Elite athletes will switch between controlled processing and automatic processing based on the difference of cognitive tasks. ③ There are also "hemisphere-specific priming" characteristics in the automatic processing of elite athletes. The left hemisphere has inhibited and decreased the linguistic control activities which are related to consciousness, while the right hemisphere has significantly enhanced which are related to the motor skills.
-
1. 问题的提出
动作技能习得的过程被划分为分化、泛化、自动化三阶段[1-2]。Fitts和Posner提出的经典三阶段模型将运动技能学习分为认知阶段(cognitive stage)、连接阶段(associative stage)、自动化阶段(autonomous stage),并认为:自动化阶段的运动技能学习过程是从有意识的控制加工向无意识的自动化加工转化;经过大量练习后,运动技能达到自动化水平,意识对动作的控制作用减少到最低限度;动作执行完全由运动程序控制,受本体感受器调节,不需要特殊的注意和纠正,机体的认知资源消耗处于节省化状态[3]。动作技能自动化包括行为层面的技能表现和心理层面的认知加工,这与认知心理学中的认知加工自动化理论保持高度一致。认知加工自动化是技能自动化的内在心理基础;技能自动化是认知加工自动化的目标,也是动作技能训练要达到的结果[4]。相关研究[5]证实,精英运动员可通过反复动作技能训练,在其专长领域呈现认知加工自动化特征,表现为所需的注意资源逐渐减少或不需要消耗注意资源[6]。Shiffrin等[7]提出人类认知加工过程中的受控加工(controlled processing)和自动化加工(automaticprocessing)双重理论。Posner等[8-9]进一步提出,认知加工自动化更强调无意识,消耗很少或不消耗认知资源,自动发生且不需要认知努力。此外,从受控加工到自动化加工是一个渐进过程而非二元对立思维[10]。
运动技能学习三阶段理论与认知加工自动化理论中的受控/自动化加工双重理论分别从运动技能习得的行为层面和心理层面[4]为动作技能自动化提供了理论基础。目前在行为层面,精英运动员动作技能自动化相关研究已较为系统,但关于精英运动员专长领域认知加工自动化特征的研究仍较少,且现有研究主要以视觉刺激参与的运动专长领域认知加工自动化解读为主[11], [12]719-742,[13],尚未有无听觉刺激参与的运动专长领域认知加工自动化研究。事实上,在运动实践中依赖听觉参与的运动项目也不在少数。如在田径短跑项目中,运动员的成绩表现与对发令员的“枪声”反应有密切关联。“起跑发令”包括“各就位”“预备”和起跑鸣枪3个阶段,这些信号均是通过听觉刺激传达给运动员的。在这3个阶段中,运动员“起跑技术”的听觉认知加工自动化特征仍有待进一步探索。综上所述,本文在对前期研究进行梳理并提出理论假设的基础上,采用感知-认知范式(perceptual-cognitive paradigms)深入探讨100 m跑精英运动员在专长领域的听觉认知加工自动化特征。
1.1 认知加工自动化特征假设1:更少的认知资源消耗
精英运动员的动作技能自动化在行为层面表现为从分化、泛化到自动化的动作呈现,在认知加工层面表现为从受控加工(需要意识参与)到自动化加工(不需要意识参与)的转化。同时,基于神经效率假说(Neural Efficiency Hypothesis),精英运动员有效的皮质功能(cortical activation)表现为更小的能量消耗[14-16],与新手组相比,精英运动员专长领域的大脑皮层激活减少[14, 17-19]。综上,得出第1个理论假设:精英运动员专长领域的动作技能自动化伴随认知加工自动化,表现为认知加工“从有意识到无意识”,以及认知资源“消耗很少或不消耗”。基于自动化渐进观点,受控加工和自动化加工之间没有绝对区分,精英运动员会根据专项任务的认知特征,灵活、高效地分配注意和意识认知资源,在受控加工和自动化加工之间进行灵活、高效的转换,即不存在一个绝对的自动化阶段。
1.2 认知加工自动化特征假设2:半球专属特有启动
梳理前人研究[20]发现:在动作技能学习初期,大脑前额叶尤其是左侧颞叶激活增加;在动作技能自动化阶段,动作执行需要的认知资源较少,通常不再呈现有意识的运动言语。在认知加工自动化阶段,大脑前额叶激活减少,控制信号传递到顶叶皮质和基底节[21-22],大脑左半球的激活会逐渐减少[23],同时右半球激活会逐渐占据主导[24]。
此外,左半球额颞区参与熟练动作行为的启动,但这些区域在动作行为启动后会受到抑制,同时左半球抑制会持续到动作行为表现结束[25]。关于射箭精英运动员的研究[26]显示,精英运动员左半球的抑制作用强于运动技能水平较低的个体。这也支持了Fitts等[27]的观点:个体在达到动作技能自动化后,不需要有意识地控制任务执行。关于高尔夫球运动员的研究[28]发现,运动员左半球激活水平与动作技能表现(高尔夫推杆命中率)呈负相关。右半球激活水平与动作技能表现呈正相关,右半球中央区、右侧颞叶和右侧顶叶区域的激活与视觉空间处理、运动任务执行的整体组织有关[12]409-476,执行复杂运动任务时右半球活动增加,同时通过训练使右半球激活增加后,精英运动员技能表现会得到显著改善。Hellige[29]基于大脑半球偏侧性提出半球专属特有启动(hemisphere-specific priming)理论,指出了半球的选择性激活特征。
综上,提出第2个理论假设:精英运动员认知加工自动化呈现半球专属特有启动特征,左半球的言语活动认知资源受到抑制,同时涉及动作技能表现的右半球激活增加[30];新手处于早期运动技能习得阶段,认知加工通过言语进行组织,左半球激活显著增加。
1.3 100 m跑起跑认知加工任务专项特征
(1)“各就位”认知任务特征。“各就位”属于100 m跑起跑信号中的次要信号,该阶段不要求高度耗费认知资源,只要求相应的行为调整:100 m跑运动员走向起跑线,完全在自己的分道内和起跑线后,按照国际田联(International Association of Athletics Federations,IAAF)径赛规则要求做好准备姿势(双手和至少1个膝盖触地,两脚接触起跑器),并等待接下来的“预备”口令。基于此,假设在“各就位”阶段:精英运动员专长领域听觉认知自动化表现出高效性和灵活性特征;而新手在该领域听觉认知加工的高效性和灵活性显著低于精英运动员,可能会耗费更多的认知资源。
(2)“预备”认知任务特征。与“各就位”信号较少消耗认知资源相比,“预备”信号属于认知资源消耗较多的警觉注意加工信号,运动员高度警觉注意即将到来的起跑鸣枪信号。假设在“预备”阶段:精英运动员在自动化认知加工阶段,随着动作技能发展呈现认知加工自动化,其前额叶激活减少,控制信号传递到顶叶皮质,同时认知加工自动化表现为左半球有意识的言语控制活动受到抑制,涉及动作技能表现的右半球激活显著增加[31];新手在认知加工初期,动作技能通过言语呈现有意识地加工[31],前额叶激活增加[21],同时涉及语言分析区域的左颞叶也参与其中[32],明确的任务表现言语(预备口令)使涉及动作技能呈现和言语加工的区域都有强烈的共同激活。
(3)起跑鸣枪认知任务特征。起跑鸣枪后的认知加工是目标探测和动作执行控制过程。个体需要高效捕获鸣枪信号,并进行快速动作执行。运动员要在瞬时产生速度和频率突增的动作执行。起跑鸣枪阶段的精英运动员专长领域认知过程自动化特征表现为能高效调动认知资源。认知加工自动化对关键刺激(起跑鸣枪)能高效调动认知资源,反之,控制加工对关键刺激(起跑鸣枪)则不能高效调动认知资源。
1.4 感知-认知范式
本文基于认知加工自动化理论以及起跑三阶段的不同认知任务,探讨精英运动员专长领域的听觉认知加工自动化特征。采用事件相关电位(Event-Related Potential,ERP)技术采集精英运动员组和普通组起跑三阶段的皮层诱发电位信号加工特征,通过专家-新手范式,寻找ERP证据并验证相关理论假设。①在“各就位”阶段,重点关注精英运动员专长领域认知资源分配;②在“预备”阶段,重点关注认知加工自动化中的左右半球专属特有启动特征;③在起跑鸣枪阶段,重点关注动作执行过程中听觉认知加工自动化调动认知资源的高效性特征。本文假设:不同认知任务的起跑三阶段脑电(electroencephalogram,EEG)活动变化存在差异,但三阶段都能体现出精英运动员认知加工自动化的专项特征,即加工过程呈现无意识或更少意识活动,很少消耗或不消耗认知资源。
Woods等[33]提出了感知-认知范式,并证实了精英运动员视觉信息加工的专家优势。基于感知-认知范式的研究[34]证实,精英运动员在特定运动中的注意力分配、物体识别和回忆、时空遮挡、视觉搜索、预测决策、执行控制等方面的表现优于新手。Woods等[33]提出,运动场景中的感知-认知技能与成功的运动表现一样重要。通过fMRI技术对比精英运动员与普通人发现,在被动接受专长领域的听觉输入时,精英运动员组脑区活动与普通组差异显著[35-38]。然而,较少有研究探讨精英运动员专长领域听觉加工自动化的中枢神经活动特征。
本文分析的是动作技能自动化的中枢神经活动(中枢神经活动控制外周运动神经[39],通过运动终板将神经冲动传递至肌纤维产生动作技能)特征,因此只要求被试听声音,不要求进行按键反应。Calvo-Merino等[40]发现,在运动场景的简单刺激输入(个体不需要动作技能呈现)过程中,运动员和普通群体呈现出认知加工资源消耗的显著差异。此外,脑机接口研究领域中关于运动表象脑神经机制的ERP研究[41-43]也要求被试只进行运动表象,不做任何按键反应。
2. 研究对象与方法
2.1 实验被试
被试由16名重庆竞技体育训练中心现役短跑运动员和17名普通人组成。运动员组被试平均训练10年以上,运动等级为国际级运动健将、运动健将或一级运动员,平均年龄为21.8岁(SD=3.60);普通组被试有100 m跑练习经验,熟悉100 m起跑三阶段口令及任务,平均年龄为21.5岁(SD=3.20)。所有被试均为右利手,无精神病史,视力或矫正视力、听力正常。实验前所有被试均被告知实验目的,签署知情同意书,并在实验后获得一定报酬。本实验已通过西南大学伦理委员会批准。
2.2 实验材料
实验语音材料采用由IAAF认证的菲普莱(Fairplay)起跑电子发令系统中的100 m跑起跑口令(“各就位”、“预备”、起跑鸣枪)。实验中,被试须佩戴入耳式耳机,在屏蔽隔音房间中记录起跑信号的听觉诱发电位。
2.3 实验设计
采用单因素(动作技能训练)被试间(运动员组-训练vs普通组-未训练)设计。运动员组经过长期短跑听口令起跑训练,普通组熟悉起跑三阶段口令及任务,但未经过长期训练。
2.4 实验程序
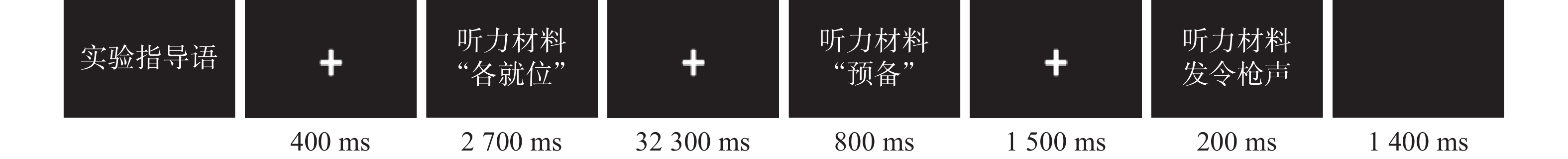
在实验过程中,被试在屏蔽隔音房间中面对一台15英寸液晶电脑显示器,接受听觉任务。在每次任务中,被试被告知全力关注起跑听觉信号。按照100 m跑起跑口令顺序呈现音频材料,根据2020年东京奥运会100 m跑半决赛和决赛起跑口令之间的平均间隔时长,设计各口令之间的时间间隔。
指导语呈现完毕后,依次呈现400 ms的注视点、“各就位”音频口令(时长2700 ms)、32300 ms的注视点,“预备”音频口令(时长800 ms)、1500 ms的注视点、起跑鸣枪信号(时长200 ms)、1400 ms的空屏。在练习实验阶段,要求被试先进行练习试次,待熟悉具体实验程序后,进入正式实验阶段。实验共72个试次,每24个试次有一个间歇(休息10 min)。具体如图1所示。
2.5 EEG数据采集与分析
2.5.1 数据采集
实验程序由E-Prime 2.0编制,实验材料的呈现由电脑控制。采用博睿康(Neuracle)公司生产的32导便携无线ERP采集系统,以Fz作为在线参考电极,记录被试对起跑信号的EEG活动情况。滤波带通为0.05~100 Hz,采样频率为300 Hz/导。头皮与电极间阻抗小于10 kΩ,采用独立成分分析(ICA),分离眼电、肌电成分去除伪迹,同时排除±80 μV的其他电极伪迹信号。
2.5.2 数据分析
完成实验记录EEG后离线处理数据,主要步骤包括:①载入数据;②以双耳乳突的平均电位为参考;③滤波为1~45 Hz;④电极定位;⑤通过独立成分分析法矫正眼电伪迹。采用EEGLAB对EEG数据进行离线数据处理;统计分析采用IBM SPSS 22.0软件。描述性统计量涉及平均波幅和标准差,同时在描述性统计的基础上做进一步方差分析。根据Luck[44]的观点,为避免对每个电极进行t检验增大统计学中的Ⅰ类错误(错误拒绝零假设)概率,将电极设为1个因素,进行组别×电极的两因素方差分析。本文主要考察个体对专长领域听觉信号的自动化加工特征,因此,ERP统计分析主要涉及听觉ERP成分。分析的ERP成分主要包括N1、P1、P2,采用基线、平均振幅进行度量。根据脑地形图、波形图以及相关文献选取电极点和成分分析的时间窗[45-47]。
3. 起跑三阶段听觉诱发电位的ERP研究结果
分析起跑三阶段数据后,剔除5名数据伪迹过多的被试,28 名被试进入最终分析,15名运动员平均年龄为22.1岁(SD=3.30),13名普通人平均年龄为21.9岁(SD=2.70)。ERP成分分析将电极设为1个因素,采用组别×电极的两因素方差分析,组别为被试间变量,电极为被试内变量。
3.1 “各就位”阶段ERP研究结果
3.1.1 ERP分析时程截取
“各就位”阶段滤波为1~45 Hz,时间窗为−200~5000 ms,基线为−200~0 ms。分析时程截取“各就位”听觉刺激呈现前200 ms至开始呈现后(包括音频持续阶段的2700 ms)的5000 ms,并以音频呈现前200 ms至音频呈现初始阶段作为基线。
3.1.2 ERP成分方差分析结果
对P2成分进行2(组别:运动员组vs普通组)×8(电极点:T6、 Fc1、Fc2、 F3、 F4、 Fz、 Fp1、 Fp2)两因素方差分析;对P3成分进行2(组别:运动员组vs普通组)×5(电极点:T5、T6、PO3、O1、OZ)两因素方差分析。各成分分析及时间窗划分如表1所示。
表 1 “各就位”阶段ERP各成分分析及时间窗划分Table 1. Component analysis of ERP on the stage of On Your Marks成分 P1 N1 P2 N2 P3 N3 P4 时间窗/ms 0~60 80~140 170~220 230~280 280~340 380~510 510~720 峰潜伏期/ms 40 115 195 255 340 430 650 差异电极 — — T6、Fc1、Fc2、F3、F4、Fz、Fp1、Fp2 T5、T6、PO3、O1 T5、T6、PO3、O1、Oz O1 — 注:“—”表示无对应差异电极。 在“各就位”阶段,时间窗为0~5000 ms,根据自动化加工实验数据处理结果、ERP成分出现的大脑区域,以及该区域中的电极点,结合相关文献研究结果,选取代表性电极,并观察其ERP总平均波形图以进行后续分析。选取的电极点如表1所示,包括:前额叶Fp1、Fp2,额叶F3、Fz、F4,额中央区Fc1、Fc2,后颞叶T5、T6,顶枕叶PO3,枕叶O1、Oz。对P2、P3成分平均振幅做进一步方差分析,同时根据每个ERP成分峰值所在的潜伏期确定时间窗,各ERP成分的波形、潜伏期不同导致对应时间窗的长度也不同。为避免1个时间窗涉及多个成分从而混淆实验结果,时间窗不宜过长,因此时间窗不连续。
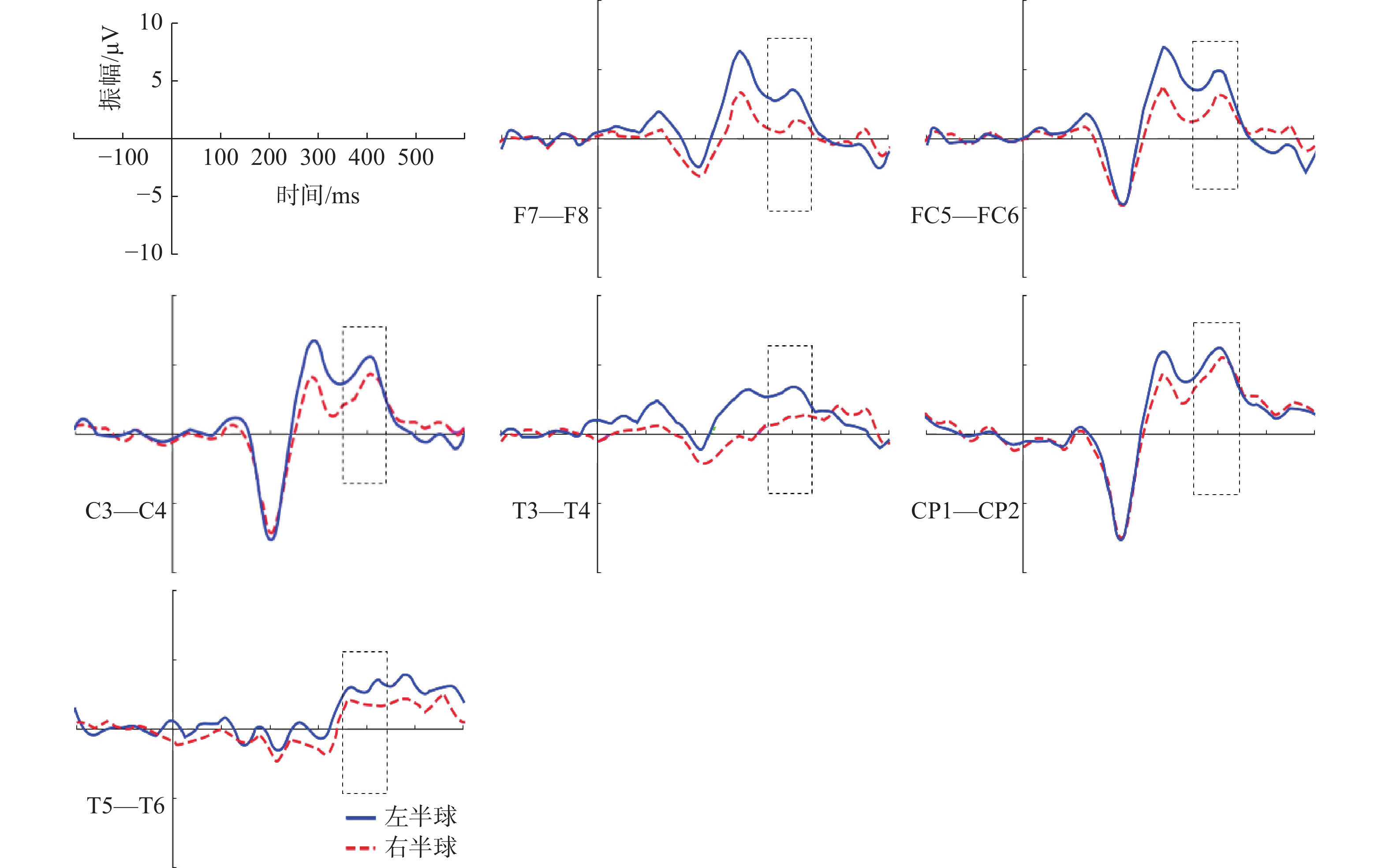
P2和P3成分选取的时间窗和对应的电极点如表1所示。对“各就位”阶段P2成分进行方差分析,采用Greenhouse-Geisser进行结果校正后得出,组别主效应显著F(1,27)=9.075,P=0.006<0.05,
$ \eta $ 2=0.259;对P3成分进行方差分析,采用Greenhouse-Geisser进行结果校正后得出,组别主效应显著F(1,27)=9.690,P=−0.004<0.05,$ \eta $ 2=0.272。P2、P3成分的波形图分别如图2、图3所示。![]() 图 2 “各就位”阶段不同组别诱发的P2波形图Figure 2. P2 components induced by different groups on the stage of On Your Marks
图 2 “各就位”阶段不同组别诱发的P2波形图Figure 2. P2 components induced by different groups on the stage of On Your Marks![]() 图 3 “各就位”阶段不同组别诱发的P3波形图Figure 3. P3 components induced by different groups on the stage of On Your Marks
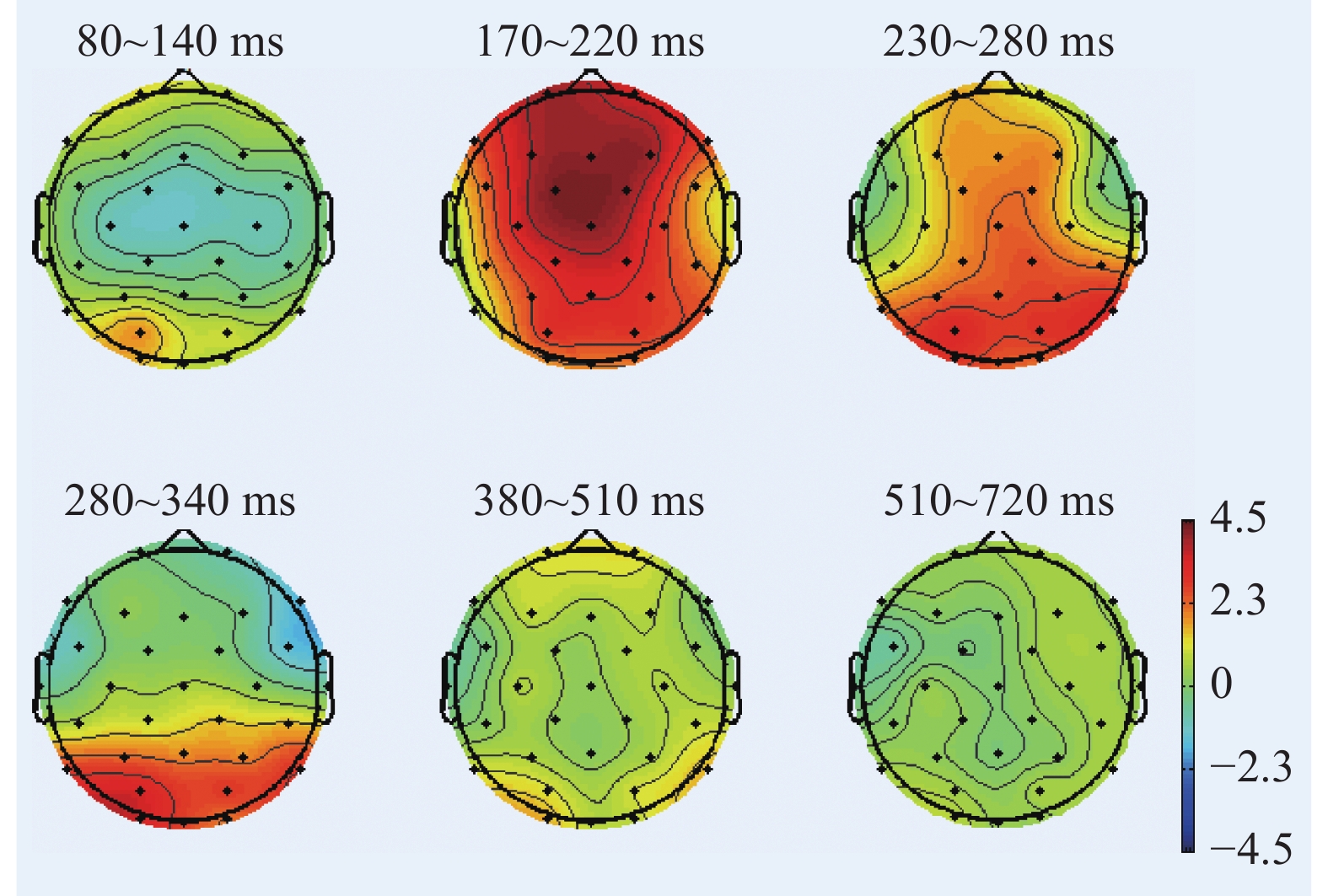
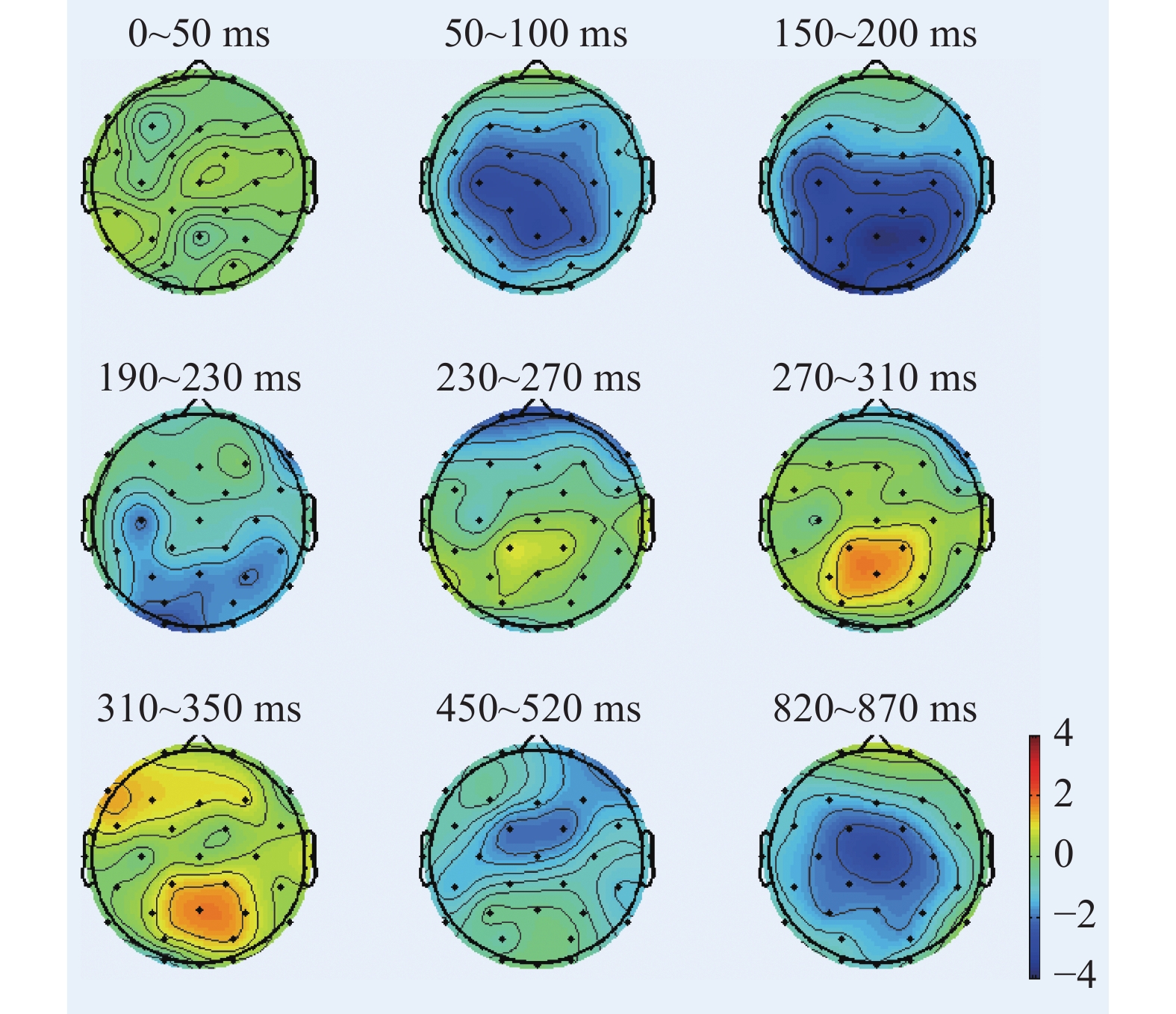
图 3 “各就位”阶段不同组别诱发的P3波形图Figure 3. P3 components induced by different groups on the stage of On Your Marks根据图2、图3绘制各时间窗内平均振幅差异脑地形图,如图4所示。在“各就位”阶段,运动员组和普通组在大脑前额叶、额叶的皮层活动差异显著,运动员组的脑激活程度显著低于普通组。
![]() 图 4 “各就位”阶段各时间窗内平均振幅差异脑地形图Figure 4. Brain topographic maps of mean amplitude difference between the two groups in each time stage (on the stage of On Your Marks)
图 4 “各就位”阶段各时间窗内平均振幅差异脑地形图Figure 4. Brain topographic maps of mean amplitude difference between the two groups in each time stage (on the stage of On Your Marks)3.2 “预备”阶段ERP研究结果
3.2.1 ERP分析时程截取
“预备”阶段滤波为1~45 Hz,时间窗为−200~4000 ms,基线为−200~0 ms。分析时程截取“预备”听觉刺激呈现前200 ms至开始呈现后(包括音频持续阶段的800 ms)的4000 ms,并以音频呈现前200 ms至音频呈现初始阶段作为基线。
3.2.2 ERP成分方差分析结果
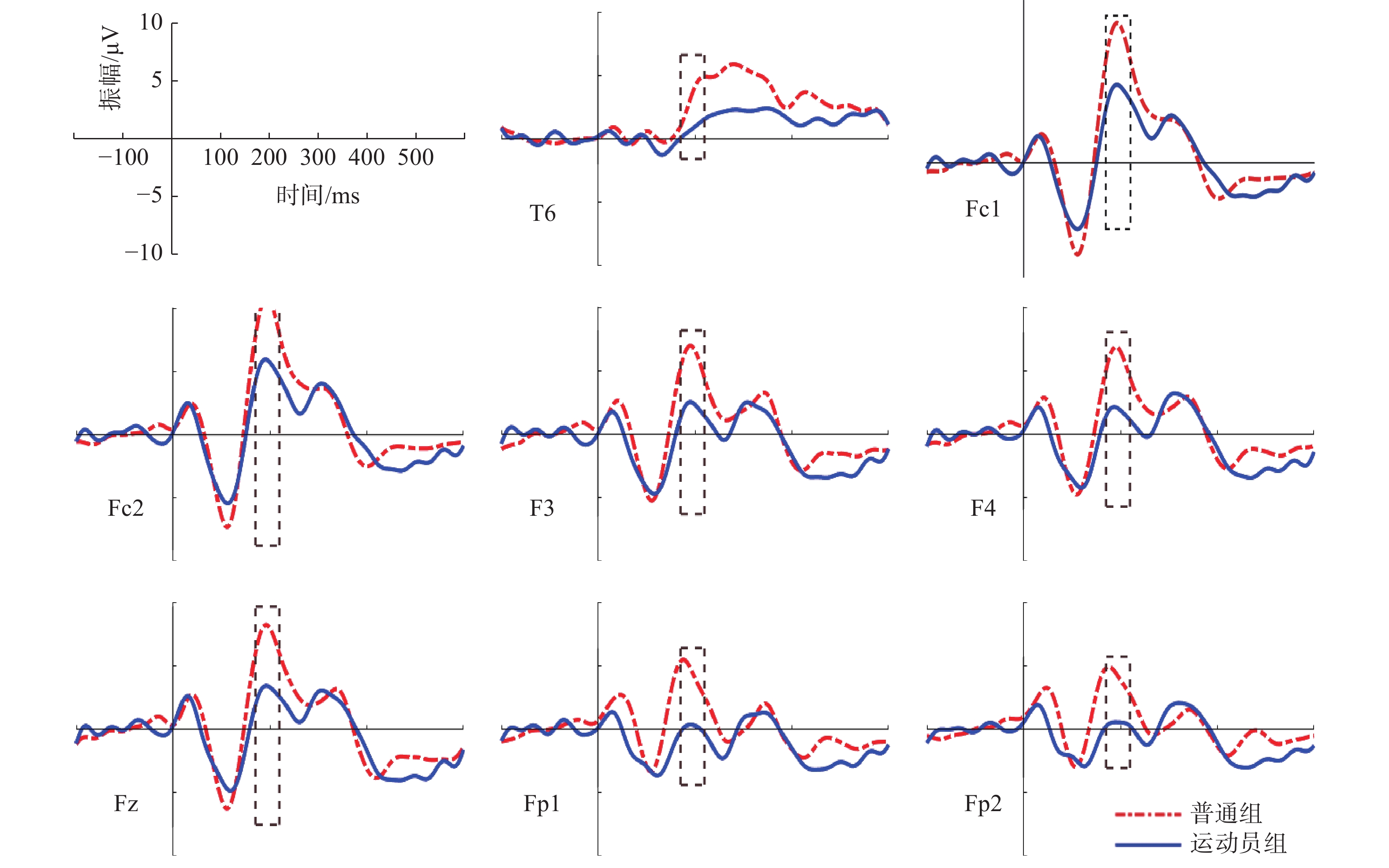
对P3成分进行2(组别:运动员组vs普通组)×8(电极点:Fz、F3、F4、F7、Fc1、Fc5、C3、T3)两因素方差分析,各成分分析及时间窗划分如表2所示。在“预备”阶段,时间窗为0~4000 ms,选取记录的电极如表2所示。
表 2 “预备”阶段ERP各成分分析及时间窗划分Table 2. Component analysis of ERP on the stage of Set成分 P1 N1 P2 N2 P3 N3 P4 时间窗/ms 80~160 180~220 270~310 310~350 350~440 440~600 600~750 峰潜伏期/ms 130 200 300 340 400 550 550 差异电极 — — F7 F7 Fz、F3、F4、F7、
Fc1、 Fc5、C3、T3T3、T5、O1 Cz、C3、C4、CP5、Pz、
P3、P4、T5、PO3、O1注:“—”表示无对应差异电极。 运动员组和普通组在电极点F7(左侧额叶)的P2波幅差异显著,P2成分波形图如图5所示。P3成分选取的时间窗为350~440 ms,分析的电极点包括额叶F7、Fz、F3、F4,额中央区Fc1、Fc5,中央区C3,颞叶T3(表2)。对“预备”阶段的P3成分进行方差分析,采用Greenhouse-Geisser进行结果校正后得出,被试间的组别效应显著F(1,29)=9.049,P=0.006<0.05,
$ \eta^2$ =0.244。P3成分波形图如图6所示。![]() 图 5 “预备”阶段不同组别诱发的P2波形图Figure 5. P2 components induced by different groups on the stage of Set
图 5 “预备”阶段不同组别诱发的P2波形图Figure 5. P2 components induced by different groups on the stage of Set![]() 图 6 “预备”阶段不同组别诱发的P3波形图Figure 6. P3 components induced by different groups on the stage of Set
图 6 “预备”阶段不同组别诱发的P3波形图Figure 6. P3 components induced by different groups on the stage of Set根据图5、图6绘制各时间窗内平均振幅差异脑地形图,如图7所示。在“预备”阶段,运动员和普通人在大脑左半球的不对称活动差异显著,同时右半球也存在一定加工能力,运动员组和普通组在左侧额叶(F7、F3、Fz)、左侧额中央(Fc1、Fc5)、左侧中央区(C3)、左颞叶(T3)、右额叶等脑区半球活动差异显著,运动员组脑激活水平显著低于普通组。
![]() 图 7 “预备”阶段各时间窗内平均振幅差异脑地形图Figure 7. Brain topographic maps of mean amplitude difference in each time stage (on the stage of Set)
图 7 “预备”阶段各时间窗内平均振幅差异脑地形图Figure 7. Brain topographic maps of mean amplitude difference in each time stage (on the stage of Set)3.2.3 左右半球专属特有启动的差异波分析
比较普通组和运动员组的P3左右半球差异波,发现两组被试的P3成分在左半球的偏侧化活动差异显著(图7)。本文分析的P3成分差异波对应的电极点有13对:Fp1—Fp2、F3—F4、F7—F8、Fc1—Fc2、Fc5—Fc6、C3—C4、T3—T4、CP1—CP2、CP5—CP6、P3—P4、T5—T6、PO3—PO4、O1—O2。普通组配对样本t检验结果显示,在F7—F8、Fc5—Fc6、C3—C4、T3—T4、CP1—CP2、T5—T6这6对电极上,出现左半球偏侧化优势(图8),左半球的平均振幅显著大于右半球。运动员组配对样本t检验结果显示,O1—O2出现右半球偏侧化优势(图9),左半球的平均振幅显著小于右半球。
![]() 图 8 普通组“预备”阶段P3成分左右半球差异Figure 8. Analysis of left and right hemispheric differences of P3 components in the ordinary group on the stage of Set
图 8 普通组“预备”阶段P3成分左右半球差异Figure 8. Analysis of left and right hemispheric differences of P3 components in the ordinary group on the stage of Set![]() 图 9 运动员组“预备”阶段P3成分左右半球差异Figure 9. Analysis of left and right hemispheric differences of P3 components in the elite group on the stage of Set
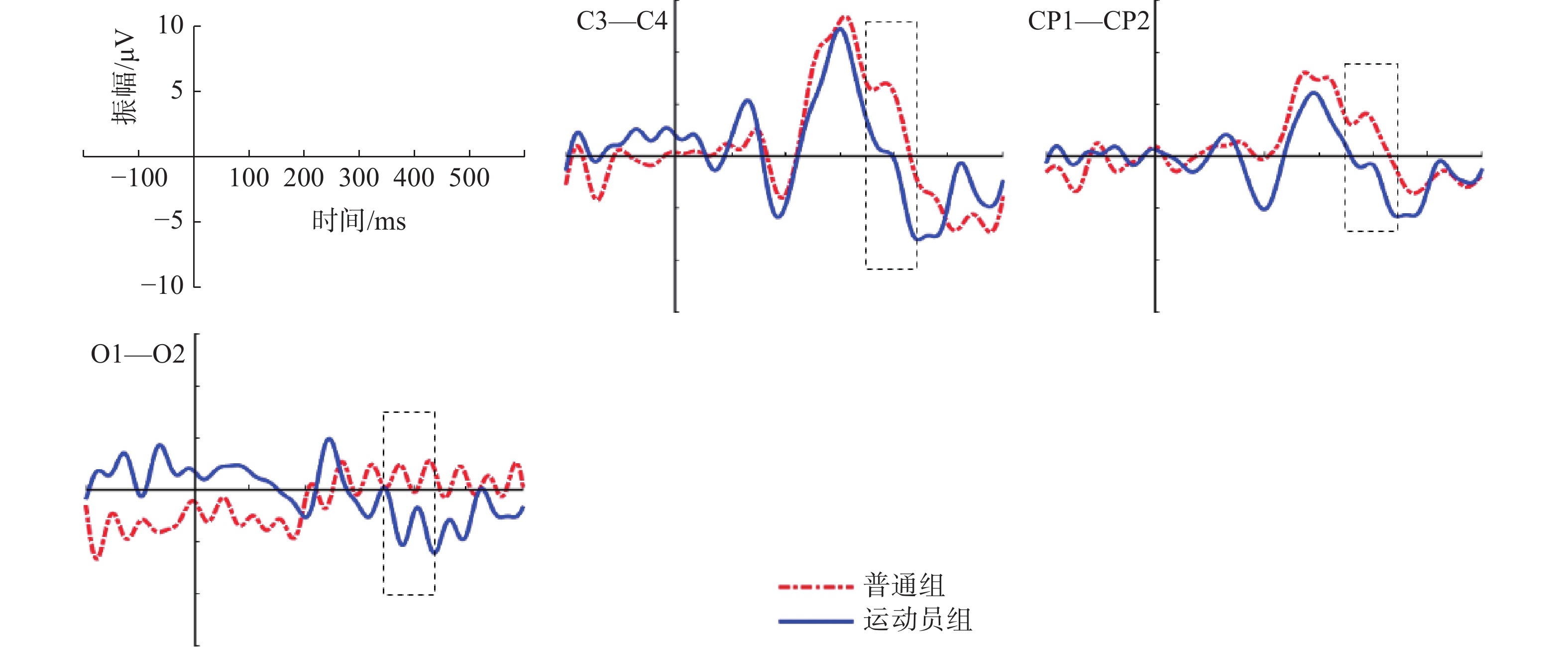
图 9 运动员组“预备”阶段P3成分左右半球差异Figure 9. Analysis of left and right hemispheric differences of P3 components in the elite group on the stage of Set将普通组和运动员组的3组差异波(C3—C4、CP1—CP2、O1—O2)进行组别×电极的两因素方差分析,采用Greenhouse-Geisser进行结果校正后得出,组别主效应显著,F(1,28)=13.719,P=0.001<0.01,
$ \eta $ 2=0.329。如图10所示:两组被试在上述3组的差异波呈现半球偏侧化优势;普通组在C3—C4、CP1—CP2的差异波呈现左半球偏侧化优势,运动员组在O1—O2的差异波呈现右半球偏侧优势。3.3 起跑鸣枪阶段ERP研究结果
3.3.1 ERP分析时程截取
起跑鸣枪阶段滤波为1~45 Hz,时间窗为−100~2000 ms,基线为−100~0 ms。分析时程截取起跑鸣枪听觉刺激呈现前100 ms至开始呈现后(包括起跑鸣枪音频持续阶段的200 ms)的2000 ms,并以音频呈现前100 ms至音频呈现初始阶段作为基线。
3.3.2 ERP成分方差分析结果
对N1成分进行2(组别:运动员组vs普通组)×18(电极点:Fc1、Fc2、Fc6、Cz、C3、C4、T4、CP1、CP2、CP5、CP6、Pz、P3、P4、T6、PO3、O1、O2)两因素方差分析;对P2成分进行2(组别:运动员组vs普通组)×11(电极点:CP5、CP6、Pz、P3、P4、T5、PO3、PO4、Oz、O1、O2)两因素方差分析。各成分分析及时间窗划分如表3所示。
表 3 起跑鸣枪阶段ERP各成分分析及时间窗划分Table 3. Component analysis of ERP on the stage of Go成分 P1 N1 P2 N2 P3 N3 P4 N4 P850 时间窗/ms 0~50 50~140 150~200 190~230 230~270 270~310 310~350 450~520 820~870 峰潜伏期/ms 15 100 170 210 250 290 330 490 850 差异电极 — Fc1、Fc2、Fc6、Cz、C3、C4、T4、
CP1、CP2、CP5、CP6、Pz、P3、
P4、T6、PO3、O1、O2CP5、CP6、Pz、P3、P4、T5、
PO3、PO4、Oz、O1、O2PO3、O1 — — — — FC1、FC5、Cz、
C3、T3、CP1、
CP2、CP5、P3注:“—”表示无对应差异电极。 在起跑鸣枪阶段,时间窗为0~2000 ms,选取记录的电极点如表3所示,包括额中央区Fc1、Fc2、Fc5、Fc6,中央区Cz、C3、C4,颞叶T3、T4,后颞叶T6、PO3,顶叶CP1、CP2、CP5、CP6、Pz、P3、P4,枕叶O1、O2。
N1和P2成分选取的时间窗和对应的电极点如表3所示。对起跑鸣枪阶段的N1成分进行方差分析,采用Greenhouse-Geisser进行结果校正后得出,被试间的组别主效应显著,F(1,30)=13.177,P=0.001<0.01,
$ \eta $ 2=0.312(图11);对P2成分进行方差分析,采用Greenhouse-Geisser进行结果校正后得出,被试间的组别主效应显著,F(1,30)=8.466,P=0.007<0.05,$ \eta $ 2=0.226(图12)。![]() 图 11 起跑鸣枪阶段不同组别诱发的N1波形图Figure 11. N1 components induced by different groups on the stage of Go
图 11 起跑鸣枪阶段不同组别诱发的N1波形图Figure 11. N1 components induced by different groups on the stage of Go![]() 图 12 起跑鸣枪阶段不同组别诱发的P2波形图Figure 12. P2 components induced by different groups on the stage of Go
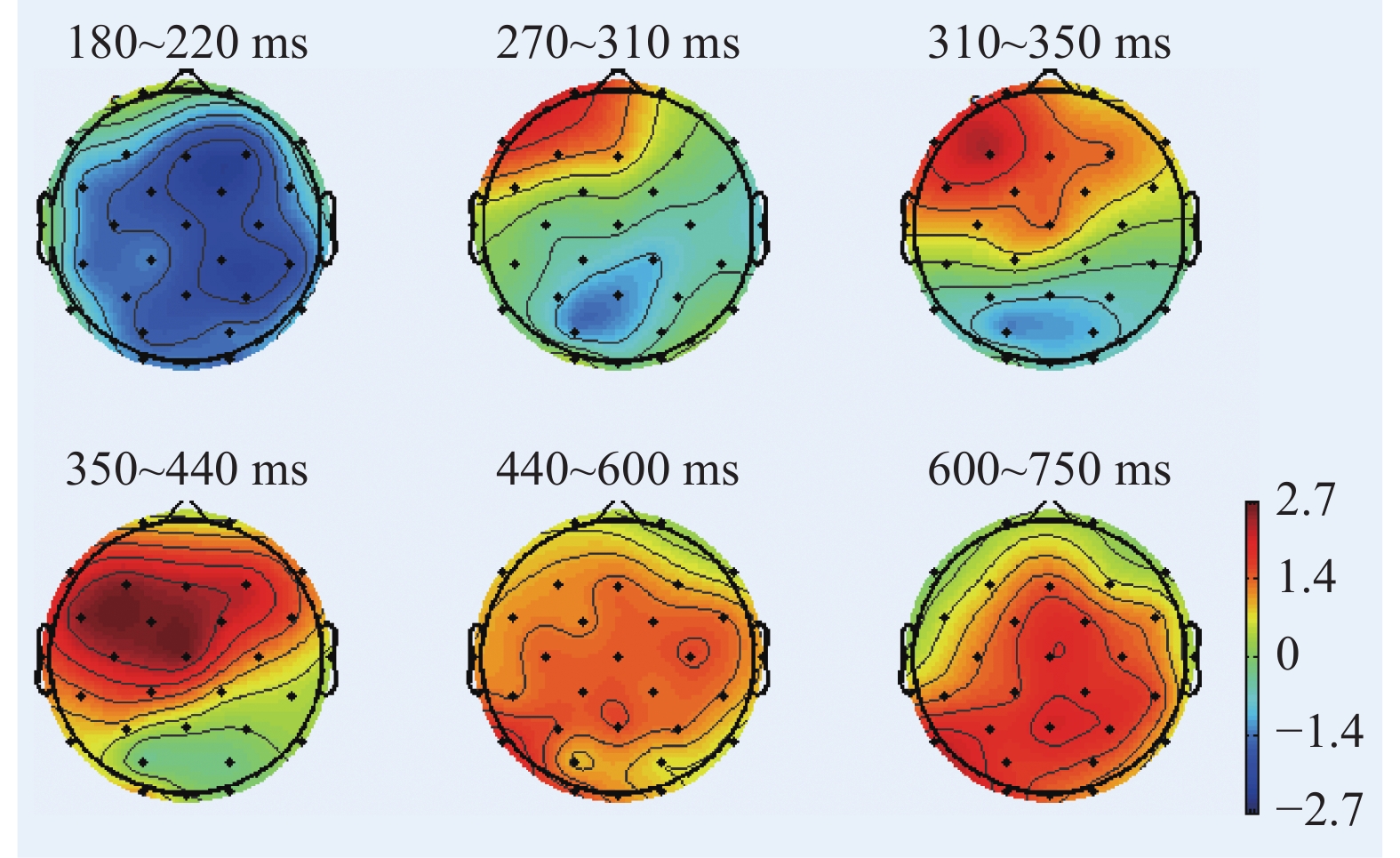
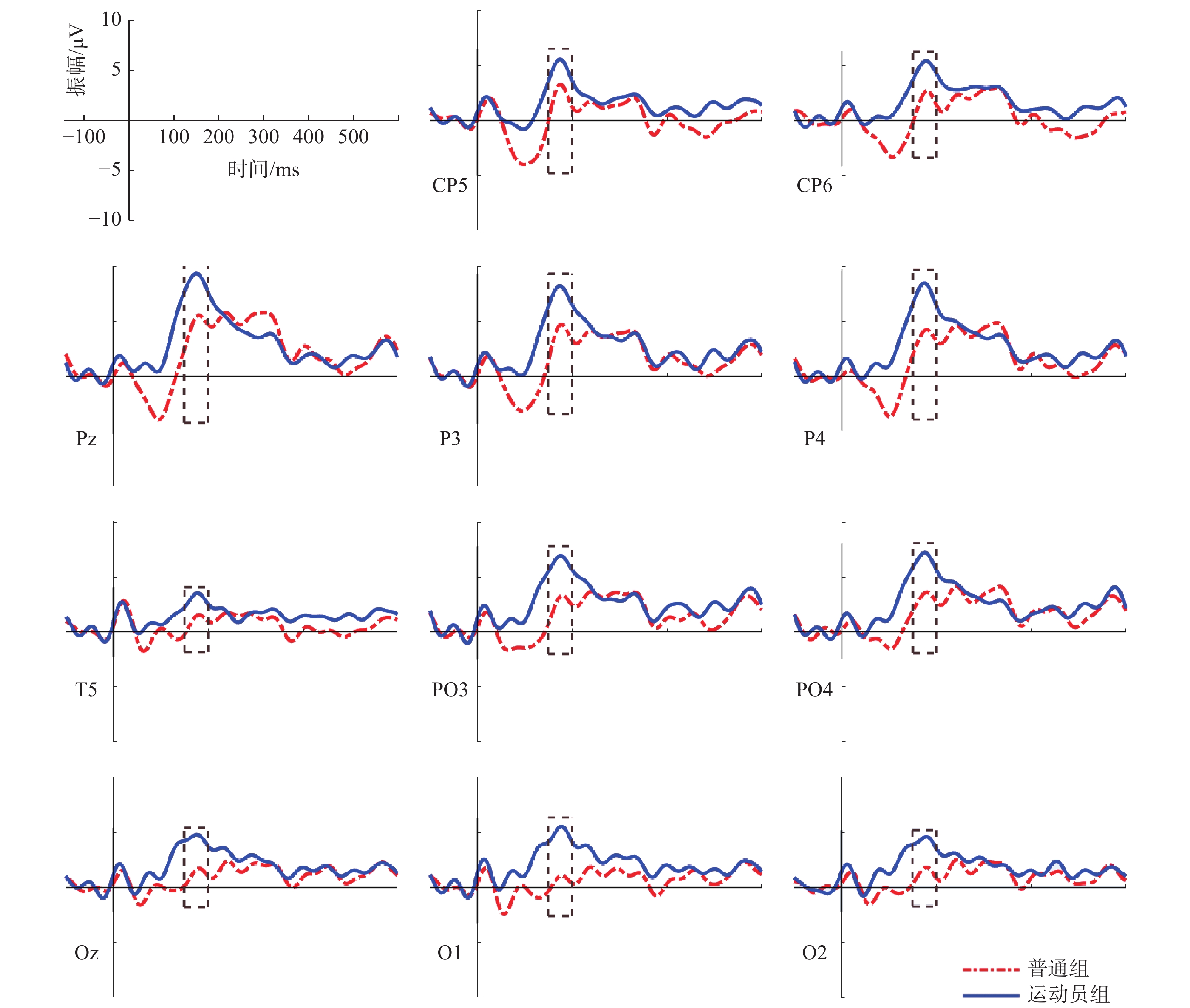
图 12 起跑鸣枪阶段不同组别诱发的P2波形图Figure 12. P2 components induced by different groups on the stage of Go根据图11、图12绘制各时间窗内平均振幅差异脑地形图,如图13所示。在起跑鸣枪阶段,运动员组和普通组在大脑额叶、颞叶、顶叶的大脑激活差异显著。运动员组N1成分的大脑激活水平显著低于普通组,P2成分的大脑激活水平显著高于普通组。
![]() 图 13 起跑鸣枪阶段各时间窗内平均振幅差异脑地形图Figure 13. Brain topographic maps of mean amplitude difference in each time stage (on the stage of Go)
图 13 起跑鸣枪阶段各时间窗内平均振幅差异脑地形图Figure 13. Brain topographic maps of mean amplitude difference in each time stage (on the stage of Go)4. 讨 论
精英运动员通过长期训练提高运用策略的能力,认知资源得到重新配置和合理使用,信息传递和处理效率显著提高[48]。此外,精英运动员可以在专长领域行为层面达到运动技能的自动化,在大脑加工活动层面呈现听觉认知加工自动化,进而产生专长领域的精英运动员认知加工自动化特征:高效解决问题,且步骤趋于自动化,基本不需要意识控制。精英运动员听觉认知加工自动化还呈现出半球专属特有启动特征,左半球涉及言语系统的皮层激活受到抑制,大脑激活水平显著降低;右半球涉及动作技能表现[49],大脑激活水平显著增强。
传统观点认为意识与注意密不可分[50-51],探寻意识的神经基础必须包含注意的作用。有研究强调,注意和意识存在重叠的神经网络,研究意识的神经基础必须包含注意[52],注意是意识的必要条件[52],其中外源性注意被认为是意识感知的先决条件[53],外源性注意捕获与后继目标的有意识感知密切相关。
本文对听觉刺激的认知加工研究主要涉及N1、P2、P3成分。N1、P2成分被认为反映了在初级听觉皮层及其附近的多个事件重叠的、空间分布的神经元对听觉刺激的预处理过程[54-55]。鉴于P2常常随着刺激与N1共同发生变化,N1—P2在部分听觉训练的研究中常被视为一个复合体,可采用峰峰振幅进行探讨(peak-to-peak amplitude)。也有部分证据显示,N1、P2之间存在部分程度的独立性[56-58],鉴于在N1没有发生显著变化的情况下,P2的功能意义可能估计不充分[45],本文独立探讨N1、P2的平均振幅。
听觉N1是刺激开始后约100 ms的波形,是一个感觉加工诱发的波形,与注意的早成分有关;也有研究认为N1反映刺激编码,N1的振幅受唤醒状态和注意的影响。Woldorff等[59]最早提出并反复验证了听觉N1的注意效应:注意可以在皮质水平加工的最初阶段影响听觉刺激加工。此外,N1可以反映对听觉刺激的意识加工[60]。P2部分反映脑干网状激活系统的听觉驱动输出[61-62],被认为可作为研究人类听觉感知训练的生理指标,经过训练后P2振幅增加,且训练对P2振幅的影响在大脑左半球呈现最大化[47]。有研究[63-64]认为,顶枕叶在刺激出现后大约200 ms呈现与意识相关的P2成分。
此外,在P3成分方面,Lumer等[65]在关于意识的研究中试图探讨意识相关神经区域(Neural Correlates of Consciousness,NCC)。Dehaene等[66]的研究进一步实证,P3振幅可以反映NCC的意识加工过程,有意识的感知会诱发顶叶和额叶的P3成分。此外,前额叶是研究意识的主要脑区之一[67],尤其是前额叶与丘脑网状核之间的注意自上而下机制,通过调节丘脑活动的能力影响意识内容的形成,且前额叶调节与意识相关的大脑交互过程。此外,Aru等[68]认为,经过训练后,对特定刺激的加工会趋于自动化,而不需要再建立工作记忆表征,使训练后的前额叶激活减少。
Posner等[69]的经典注意网络理论(authority attention network theory)认为,注意是一个复杂结构,包含多个功能和解剖上分离但相互作用的网络,具有警觉、定向和执行控制功能,额叶、顶叶、丘脑与唤醒、警觉有关,并且与对即将输入刺激的敏感度和反应准备有关。P3与警觉和执行控制有关,P3振幅的大小反映了分配给外源性注意的认知资源量[70]。运动使得额叶、中央区、顶叶中线区域的P3(涉及警觉和执行控制子任务)振幅增大[71]。研究[72-73]发现,急性运动(acute exercise)后认知效率改善,并影响P3振幅,这可能是由于分配给后续任务的认知资源在短期内得到增强所致。额顶叶网络活动既与注意有关,也与意识有关。相关研究[74]证实,外源性注意通过额顶叶网络诱发P3成分,有意识地参与感知加工,说明外源性注意是意识知觉的先决条件。
4.1 “各就位”阶段听觉认知加工自动化ERP特征
“各就位”阶段的精英运动员专长领域自动化加工在无意识中进行,自动化加工过程很少消耗或根本不消耗认知资源。本文“各就位”阶段运动员组P2成分在前额叶、额叶、额中央、右后颞叶的平均振幅显著低于普通组,运动员组P3成分平均振幅在颞叶、枕后区和枕叶显著低于普通组。运动员组在听觉意识感知、驱动输出、警觉注意、动作执行等方面分配给“各就位”信号的认知加工资源量较少,其额叶和前额叶的激活程度显著低于普通组,说明精英运动员抑制无关信息的能力比普通人更高,消耗的认知加工资源显著低于普通人。本文结果显示,运动员组在次要认知信号输入阶段,消耗的认知资源显著低于普通组,符合认知加工自动化特征假设1。
前额叶是NCC的重要组成部分,其激活水平与执行功能、自上而下或自下而上的控制有关。对刺激进行训练获得经验会导致记忆表征的建立,从而通过训练减少前额活动。本文结果与上述理论一致,运动员组前额叶的活动显著低于普通组,因此认为在“各就位”阶段认知加工自动化的特征体现为:运动员专长领域的认知资源消耗显著较少,涉及意识加工的前额叶激活显著减少。
4.2 “预备”阶段听觉认知加工自动化ERP特征
在高度警觉注意的“预备”阶段,运动员组的认知加工资源消耗量仍显著低于普通组,有意识加工使得普通组左半球活动显著增强。本文结果显示,运动员组在左半球的抑制作用显著强于普通组,而涉及动作技能表现的右半球激活差异不显著。同时,左半球涉及言语意识控制的活动显著增强,会干扰自动化认知。与之相对,运动员在警觉注意的“预备”阶段,不需要有意识的言语控制,出现听觉认知加工自动化。呈现半球专属特有启动特征:左半球涉及言语意识控制区域的激活显著减少,右半球涉及动作技能表现的活动显著增强。运动员组P2成分在左侧额叶的振幅显著低于普通组,P3成分在额叶、左侧额叶、左侧额中央、左侧中央区、左侧颞叶的振幅显著低于普通组,说明精英运动员在高度警觉状态下P3成分左半球加工偏侧性减少,抑制作用显著强于普通人。本文对“预备”阶段P3成分进行左右半球差异波分析,证实了左半球抑制能力与卓越技能表现呈正相关。同时,运动员组P3成分振幅显著低于普通组,符合自动化加工特征:意识和注意资源的消耗显著减少,且自动化发生不需要认知努力。
4.3 起跑鸣枪阶段听觉认知加工自动化ERP特征
听觉N1成分是一个感觉加工诱发的波形,同时可以反映对听觉刺激的意识加工。 Woldorff等[59]和 Hillyard等[75]的研究显示,听觉N1振幅受注意影响,注意加工水平与听觉N1振幅成正比,注意可以在皮质加工的最初阶段影响听觉刺激加工。运动员组N1成分在额中央区、中央区、中央顶叶、顶叶、 颞叶、顶枕叶区域的平均振幅显著低于普通组。精英运动员对于鸣枪信号的注意呈现无意识的自动化加工特征,说明其对专长领域的信号加工不占用注意资源,证实了精英运动员专长领域听觉加工的自动化特征。
P2成分反映意识加工,同时反映脑干网状激活系统的听觉驱动输出和动作执行控制[47]。运动员组P2成分在中央顶叶、顶叶、颞叶、顶枕叶区域的平均振幅高于普通组,表明精英运动员将总量有限的注意资源大部分调配至起跑鸣枪阶段的听觉驱动输出和动作执行的神经活动方面。在大脑皮层活动空间层面,表现为精英运动员在顶叶皮层(顶叶皮层是NCC的必要先决条件[67],同时涉及听觉注意执行控制)和脑干区域(听觉驱动输出)的P2振幅显著高于普通人。在起跑鸣枪阶段,运动员组消耗的注意和意识资源显著高于普通组,支持了当前认知加工自动化理论中的渐进观,即不存在一个绝对自动化的阶段,自动化不是“全或无”,受控加工也参与自动化加工的启动阶段[76]。精英运动员在专长领域处理认知信号时,也会基于认知任务差异,在受控加工和自动化加工二者之间进行转换。
4.4 本文局限性
本文选取的运动员被试是国际级运动健将、运动健将或一级运动员,被试的特殊性导致样本量较少,同时运动员被试经历了10年以上甚至更长时间的训练,与仅熟悉100 m跑起跑信号的普通人群体相比,群体间差异较大。未来的研究应采用更大的样本量,这将有助于深入探究两类群体之间差异细微的神经机制。
5. 结论与展望
(1)本文基于认知加工自动化和技能习得自动化的基础理论,证实了在专长领域认知加工自动化方面,精英运动员与普通人的中枢神经活动特征差异表现为精英运动员不需要或只占用较少认知资源,所需的注意和意识资源逐渐减少。这支持了认知加工自动化的神经效率假说。
(2)精英运动员根据专项经验,高效灵活调配认知资源,证实了“条件化”自动化加工理论提出的“认知系统会根据注意或任务的不同进行调整,以保证自动化加工的顺利产生和运行”;同时,还支持“采用受控加工与意识加工的渐进观点”来解释动作技能自动化现象。
(3)在100 m跑的警觉注意“预备”阶段,精英运动员听觉认知加工自动化呈现半球专属特有启动特征:精英运动员左半球有意识的言语控制活动受到抑制,右半球涉及动作技能表现的活动显著增强。
以往研究中的运动偏侧性问题多集中于存在较多单侧性动作技能的非对称性项目(如乒乓球、棒球、击剑、柔道、摔跤、拳击等)[77-78],然而本文发现,在100 m跑的对称性运动项目的“预备”阶段,同样呈现大脑皮层活动的偏侧性(非均衡性)特征。未来的研究应进一步探讨认知加工自动化过程中偏侧性的影响因素,以及偏侧性和最佳竞技表现的关系。
作者贡献声明:朱瑜:提出论文主题,设计论文框架,撰写论文;作者贡献声明:朱琪琪:修改图表;作者贡献声明:许翀:梳理文献,采集数据;作者贡献声明:刘湘:联系被试,协助采集数据;作者贡献声明:袁宏、郭立亚:指导修改论文;作者贡献声明:王进:修改论文。 -
![]()
图 2 “各就位”阶段不同组别诱发的P2波形图
Figure 2. P2 components induced by different groups on the stage of On Your Marks
![]()
图 3 “各就位”阶段不同组别诱发的P3波形图
Figure 3. P3 components induced by different groups on the stage of On Your Marks
![]()
图 4 “各就位”阶段各时间窗内平均振幅差异脑地形图
Figure 4. Brain topographic maps of mean amplitude difference between the two groups in each time stage (on the stage of On Your Marks)
![]()
图 5 “预备”阶段不同组别诱发的P2波形图
Figure 5. P2 components induced by different groups on the stage of Set
![]()
图 6 “预备”阶段不同组别诱发的P3波形图
Figure 6. P3 components induced by different groups on the stage of Set
![]()
图 7 “预备”阶段各时间窗内平均振幅差异脑地形图
Figure 7. Brain topographic maps of mean amplitude difference in each time stage (on the stage of Set)
![]()
图 8 普通组“预备”阶段P3成分左右半球差异
Figure 8. Analysis of left and right hemispheric differences of P3 components in the ordinary group on the stage of Set
![]()
图 9 运动员组“预备”阶段P3成分左右半球差异
Figure 9. Analysis of left and right hemispheric differences of P3 components in the elite group on the stage of Set
![]()
图 11 起跑鸣枪阶段不同组别诱发的N1波形图
Figure 11. N1 components induced by different groups on the stage of Go
![]()
图 12 起跑鸣枪阶段不同组别诱发的P2波形图
Figure 12. P2 components induced by different groups on the stage of Go
![]()
图 13 起跑鸣枪阶段各时间窗内平均振幅差异脑地形图
Figure 13. Brain topographic maps of mean amplitude difference in each time stage (on the stage of Go)
表 1 “各就位”阶段ERP各成分分析及时间窗划分
Table 1 Component analysis of ERP on the stage of On Your Marks
成分 P1 N1 P2 N2 P3 N3 P4 时间窗/ms 0~60 80~140 170~220 230~280 280~340 380~510 510~720 峰潜伏期/ms 40 115 195 255 340 430 650 差异电极 — — T6、Fc1、Fc2、F3、F4、Fz、Fp1、Fp2 T5、T6、PO3、O1 T5、T6、PO3、O1、Oz O1 — 注:“—”表示无对应差异电极。  下载: 导出CSV
下载: 导出CSV
表 2 “预备”阶段ERP各成分分析及时间窗划分
Table 2 Component analysis of ERP on the stage of Set
成分 P1 N1 P2 N2 P3 N3 P4 时间窗/ms 80~160 180~220 270~310 310~350 350~440 440~600 600~750 峰潜伏期/ms 130 200 300 340 400 550 550 差异电极 — — F7 F7 Fz、F3、F4、F7、
Fc1、 Fc5、C3、T3T3、T5、O1 Cz、C3、C4、CP5、Pz、
P3、P4、T5、PO3、O1注:“—”表示无对应差异电极。
下载: 导出CSV
表 3 起跑鸣枪阶段ERP各成分分析及时间窗划分
Table 3 Component analysis of ERP on the stage of Go
成分 P1 N1 P2 N2 P3 N3 P4 N4 P850 时间窗/ms 0~50 50~140 150~200 190~230 230~270 270~310 310~350 450~520 820~870 峰潜伏期/ms 15 100 170 210 250 290 330 490 850 差异电极 — Fc1、Fc2、Fc6、Cz、C3、C4、T4、
CP1、CP2、CP5、CP6、Pz、P3、
P4、T6、PO3、O1、O2CP5、CP6、Pz、P3、P4、T5、
PO3、PO4、Oz、O1、O2PO3、O1 — — — — FC1、FC5、Cz、
C3、T3、CP1、
CP2、CP5、P3注:“—”表示无对应差异电极。
下载: 导出CSV
-
[1] 邓树勋, 王健, 乔德才. 运动生理学[M]. 3版. 北京: 高等教育出版社, 2015: 285-293 [2] 张力为, 毛志雄. 运动心理学[M]. 上海: 华东师范大学出版社, 2003: 215-220 [3] 胡桂英. 运动技能内隐学习的特征、机制和应用研究[D]. 杭州: 浙江大学, 2009: 7-8 [4] 陈圣栋,陈永强,高伟,等. 认知加工的自动化现象:从二分法到渐进观[J]. 心理科学进展,2019,27(9):1556-1563 [5] STERNBERG R J, STERNBERG K.认知心理学[M].6版. 邵志芳, 译. 北京: 中国轻工业出版社, 2016: 152-154 [6] MOORS A,DE HOUWER J. Automaticity:A theoretical and conceptual analysis[J]. Psychological Bulletin,2006,132(2):297-326 doi: 10.1037/0033-2909.132.2.297
[7] SHIFFRIN R M,SCHNEIDER W. Controlled and automatic human information processing:Ⅱ. Perceptual learning,automatic attending and a general theory[J]. Psychological Review,1977,84(2):127-190 doi: 10.1037/0033-295X.84.2.127
[8] POSNER M I, SNYDER C R R. Facilitation and inhibition in the processing of signals[M]//RABBITT P M A, DORNIC S. Attention and performance. New York: Academic Press, 1975: 669–682
[9] POSNER M I, SNYDER C R R. Attention and cognitive control[M]//BALOTA D A, MARSH E J. Cognitive psychology: Key readings. New York, NY: Psychology Press, 2004: 205-223
[10] BARGH J A. The ecology of automaticity:Toward establishing the conditions needed to produce automatic processing effects[J]. The American Journal of Psychology,1992,105(2):181-199 doi: 10.2307/1423027
[11] HU Y,ERICSSON K A,YANG D,et al. Superior self-paced memorization of digits in spite of a normal digit span:The structure of a memorist's skill[J]. Journal of Experimental Psychology. Learning,Memory,and Cognition,2009,35(6):1426-1442 doi: 10.1037/a0017395
[12] HUGDAHL K, DAVIDSON R J. The asymmetrical brain[M]. Cambridge, Mass. : MIT Press, 2003
[13] BLAXTON T A. Distinguishing false from true in human memory[J]. Neuron,1996,17(2):191-194 doi: 10.1016/S0896-6273(00)80150-6
[14] PERCIO C D,INFARINATO F,IACOBONI M,et al. Movement-related desynchronization of alpha rhythms is lower in athletes than non-athletes:A high-resolution EEG study[J]. Clinical Neurophysiology,2010,121(4):482-491 doi: 10.1016/j.clinph.2009.12.004
[15] 王进. “神经效率”假说在体育中的验证探索[J]. 体育科学,2009,29(4):97 doi: 10.16469/j.css.2009.04.014 [16] 闫鹏,刘文学,赵怀阳. 运动员神经效率假说的相关研究综述[J]. 科学中国人,2014(22):159 [17] BABILONI C,MARZANO N,INFARINATO F,et al. "Neural efficiency" of experts' brain during judgment of actions:A high-resolution EEG study in elite and amateur karate athletes[J]. Behavioural Brain Research,2010,207(2):466-475 doi: 10.1016/j.bbr.2009.10.034
[18] 郭志平. 乒乓球运动员在视觉—空间认知加工中的神经效率及其神经机制[D]. 上海: 上海体育学院, 2016: 86 [19] 魏瑶. 乒乓球运动员在不同空间任务认知加工神经效率的EEG研究[D]. 上海: 上海体育学院, 2019: 103-105 [20] LACOURSE M G,ORR E L R,CRAMER S C,et al. Brain activation during execution and motor imagery of novel and skilled sequential hand movements[J]. NeuroImage,2005,27(3):505-519 doi: 10.1016/j.neuroimage.2005.04.025
[21] DOYON J,PENHUNE V,UNGERLEIDER L G. Distinct contribution of the cortico-striatal and cortico-cerebellar systems to motor skill learning[J]. Neuropsychologia,2003,41(3):252-262 doi: 10.1016/S0028-3932(02)00158-6
[22] VAN MIER H,TEMPEL L W,PERLMUTTER J S,et al. Changes in brain activity during motor learning measured with PET:Effects of hand of performance and practice[J]. Journal of Neurophysiology,1998,80(4):2177-2199 doi: 10.1152/jn.1998.80.4.2177
[23] LANDERS D,HAN M,SALAZAR W,et al. Effects of learning on electroencephalographic and electrocardiographic patterns in novice archers[J]. International Journal of Sport Psychology,1994,25(3):313-330
[24] SALAZAR W,LANDERS D M,PETRUZZELLO S J,et al. Hemispheric asymmetry,cardiac response,and performance in elite archers[J]. Res Q Exerc Sport,1990,61(4):351-359 doi: 10.1080/02701367.1990.10607499
[25] HATFIELD B D,HAUFLER A J,HUNG T M,et al. Electroencephalographic studies of skilled psychomotor performance[J]. Journal of Clinical Neurophysiology,2004,21(3):144-156 doi: 10.1097/00004691-200405000-00003
[26] CREWS D J,LANDERS D M. Electroencephalographic measures of attentional patterns prior to the golf putt[J]. Medicine and Science in Sports and Exercise,1993,25(1):116-126 doi: 10.1249/00005768-199301000-00016
[27] FITTS P M, POSNER M I. Human performance[M]. Belmont, CA: Brooke/Cole, 1967: 162
[28] CREWS D J. What your brain is doing when you putt[J]. Golf Digest,2004,1:100-101
[29] HELLIGE J B. Hemispheric asymmetry: What's right and what's left[M]. Cambridge, Mass.: Harvard University Press, 1993: 104-109
[30] HILLMAN C H,APPARIES R J,JANELLE C M,et al. An electrocortical comparison of executed and rejected shots in skilled marksmen[J]. Biological Psychology,2000,52(1):71-83 doi: 10.1016/S0301-0511(99)00021-6
[31] BEILOCK S L,CARR T H. On the fragility of skilled performance:What governs choking under pressure?[J]. Journal of Experimental Psychology:General,2001,130(4):701-725 doi: 10.1037/0096-3445.130.4.701
[32] DEENY S P,HILLMAN C H,JANELLE C M,et al. Cortico-cortical communication and superior performance in skilled marksmen:An EEG coherence analysis[J]. Journal of Sport and Exercise Psychology,2003,25(2):188-204 doi: 10.1123/jsep.25.2.188
[33] WOODS E A,HERNANDEZ A E,WAGNER V E,et al. Expert athletes activate somatosensory and motor planning regions of the brain when passively listening to familiar sports sounds[J]. Brain and Cognition,2014,87:122-133 doi: 10.1016/j.bandc.2014.03.007
[34] ZHANG D D,DING H Y,WANG X C,et al. Enhanced response inhibition in experienced fencers[J]. Scientific Reports,2015,5:16282 doi: 10.1038/srep16282
[35] MALOUIN F,RICHARDS C L,JACKSON P L,et al. Brain activations during motor imagery of locomotor-related tasks:A PET study[J]. Human Brain Mapping,2003,19(1):47-62 doi: 10.1002/hbm.10103
[36] PORRO C A,CETTOLO V,FRANCESCATO M P,et al. Ipsilateral involvement of primary motor cortex during motor imagery[J]. The European Journal of Neuroscience,2000,12(8):3059-3063 doi: 10.1046/j.1460-9568.2000.00182.x
[37] SHERGILL S S,BULLMORE E T,BRAMMER M J,et al. A functional study of auditory verbal imagery[J]. Psychological Medicine,2001,31(2):241-253 doi: 10.1017/S003329170100335X
[38] WANG C H,WAI Y,WENG Y,et al. The cortical modulation from the external cues during gait observation and imagination[J]. Neuroscience Letters,2008,443(3):232-235 doi: 10.1016/j.neulet.2008.07.084
[39] 袁琼嘉, 李雪. 运动解剖学[M]. 北京: 高等教育出版社, 2019: 270-271 [40] CALVO-MERINO B,GRÈZES J,GLASER D E,et al. Seeing or doing? Influence of visual and motor familiarity in action observation[J]. Current Biology,2006,16(19):1905-1910 doi: 10.1016/j.cub.2006.07.065
[41] 张锐. 运动想象脑-机接口的神经机制与识别算法研究[D]. 成都: 电子科技大学, 2015: 18-19 [42] 施锦河. 运动想象脑电信号处理与P300刺激范式研究[D]. 杭州: 浙江大学, 2012: 8-11 [43] 周鹏. 基于运动想象的脑机接口的研究[D]. 天津: 天津大学, 2007: 11 [44] LUCK S J. 事件相关电位基础[M]. 范思陆, 译. 上海: 华东师范大学出版社, 2009: 195 [45] ANDERSON S,KRAUS N. Auditory training:Evidence for neural plasticity in older adults[J]. Perspectives on Hearing and Hearing Disorders. Research and Research Diagnostics,2013,17:37-57 doi: 10.1044/hhd17.1.37
[46] TREMBLAY K,MCCLANNAHAN K. Auditory training:Stimulus exposure,task execution,and response feedback affect the neural detection of sound[J]. The Journal of the Acoustical Society of America,2011,129(4):2660
[47] TREMBLAY K L,SHAHIN A J,PICTON T,et al. Auditory training alters the physiological detection of stimulus-specific cues in humans[J]. Clinical Neurophysiology,2009,120(1):128-135 doi: 10.1016/j.clinph.2008.10.005
[48] 祝一虹,董宗旺,翁旭初,等. 短时练习对序列运动脑功能偏侧化的影响[J]. 科学通报,2005,50(3):215-218 doi: 10.3321/j.issn:0023-074X.2005.03.004 [49] 蔡绍芃, 董国亚, 白俊. 脑功能偏侧化的研究[C]//天津市生物医学工程学会第三十一届学术年会论文集. 天津, 2011: 42 [50] O'REGAN J K,NOË A. A sensorimotor account of vision and visual consciousness[J]. The Behavioral and Brain Sciences,2001,24(5):939-973 doi: 10.1017/S0140525X01000115
[51] DE BRIGARD F,PRINZ J. Attention and consciousness[J]. WIREs Cognitive Science,2010,1(1):51-59 doi: 10.1002/wcs.27
[52] COHEN M A,CAVANAGH P,CHUN M M,et al. The attentional requirements of consciousness[J]. Trends in Cognitive Sciences,2012,16(8):411-417 doi: 10.1016/j.tics.2012.06.013
[53] CHICA A B,LASAPONARA S,LUPIÁÑEZ J,et al. Exogenous attention can capture perceptual consciousness:ERP and behavioural evidence[J]. NeuroImage,2010,51(3):1205-1212 doi: 10.1016/j.neuroimage.2010.03.002
[54] KOYAMA S,GUNJI A,YABE H,et al. Hemispheric lateralization in an analysis of speech sounds[J]. Cognitive Brain Research,2000,10(1-2):119-124 doi: 10.1016/S0926-6410(00)00034-3
[55] SHTYROV Y,KUJALA T,LYYTINEN H,et al. Auditory cortex evoked magnetic fields and lateralization of speech processing[J]. NeuroReport,2000,11(13):2893-2896 doi: 10.1097/00001756-200009110-00013
[56] GODEY B,SCHWARTZ D,DE GRAAF J B,et al. Neuromagnetic source localization of auditory evoked fields and intracerebral evoked potentials:A comparison of data in the same patients[J]. Clinical Neurophysiology,2001,112(10):1850-1859 doi: 10.1016/S1388-2457(01)00636-8
[57] HARKRIDER A W,PLYLER P N,HEDRICK M S. Effects of age and spectral shaping on perception and neural representation of stop consonant stimuli[J]. Clinical Neurophysiology,2005,116(9):2153-2164 doi: 10.1016/j.clinph.2005.05.016
[58] ROSS B,TREMBLAY K L,PICTON T W. Physiological detection of interaural phase differences[J]. The Journal of the Acoustical Society of America,2007,121(2):1017-1027 doi: 10.1121/1.2404915
[59] WOLDORFF M G,HILLYARD S A. Modulation of early auditory processing during selective listening to rapidly presented tones[J]. Electroencephalography and Clinical Neurophysiology,1991,79(3):170-191 doi: 10.1016/0013-4694(91)90136-R
[60] EDMONDS B A,JAMES R E,UTEV A,et al. Evidence for early specialized processing of speech formant information in anterior and posterior human auditory cortex[J]. The European Journal of Neuroscience,2010,32(4):684-692 doi: 10.1111/j.1460-9568.2010.07315.x
[61] RIF J,HARI R,HÄMÄLÄINEN M S,et al. Auditory attention affects two different areas in the human supratemporal cortex[J]. Electroencephalography and Clinical Neurophysiology,1991,79(6):464-472 doi: 10.1016/0013-4694(91)90166-2
[62] WOODS D L,KNIGHT R T,SCABINI D. Anatomical substrates of auditory selective attention:Behavioral and electrophysiological effects of posterior association cortex lesions[J]. Cognitive Brain Research,1993,1(4):227-240 doi: 10.1016/0926-6410(93)90007-R
[63] BUSCH N A,DUBOIS J,VANRULLEN R. The phase of ongoing EEG oscillations predicts visual perception[J]. Journal of Neuroscience,2009,29(24):7869-7876 doi: 10.1523/JNEUROSCI.0113-09.2009
[64] MATHEWSON K E,GRATTON G,FABIANI M,et al. To see or not to see:Prestimulus phase predicts visual awareness[J]. Journal of Neuroscience,2009,29(9):2725-2732 doi: 10.1523/JNEUROSCI.3963-08.2009
[65] LUMER E D,REES G. Covariation of activity in visual and prefrontal cortex associated with subjective visual perception[J]. Proceedings of the National Academy of Sciences of the United States of America,1999,96(4):1669-1673 doi: 10.1073/pnas.96.4.1669
[66] DEHAENE S,CHANGEUX J P,NACCACHE L,et al. Conscious,preconscious,and subliminal processing:A testable taxonomy[J]. Trends in Cognitive Sciences,2006,10(5):204-211 doi: 10.1016/j.tics.2006.03.007
[67] LEÓN-DOMÍNGUEZ U,LEÓN-CARRIÓN J. Prefrontal neural dynamics in consciousness[J]. Neuropsychologia,2019,131:25-41 doi: 10.1016/j.neuropsychologia.2019.05.018
[68] ARU J,BACHMANN T,SINGER W,et al. Distilling the neural correlates of consciousness[J]. Neuroscience & Biobehavioral Reviews,2012,36(2):737-746
[69] POSNER M I,PETERSEN S E. The attention system of the human brain[J]. Annual Review of Neuroscience,1990,13:25-42 doi: 10.1146/annurev.ne.13.030190.000325
[70] HILLMAN C H,KAMIJO K,SCUDDER M. A review of chronic and acute physical activity participation on neuroelectric measures of brain health and cognition during childhood[J]. Preventive Medicine,2011,52:S21-S28 doi: 10.1016/j.ypmed.2011.01.024
[71] CHANG Y K,PESCE C,CHIANG Y T,et al. Antecedent acute cycling exercise affects attention control:An ERP study using attention network test[J]. Frontiers in Human Neuroscience,2015,9:156
[72] KAMIJO K,NISHIHIRA Y,HATTA A,et al. Differential influences of comfort level obtained after acute exercise on information processing in the central nervous system[J]. Taiikugaku Kenkyu,2005,50(6):663-673
[73] KAMIJO K,NISHIHIRA Y,HIGASHIURA T,et al. The interactive effect of exercise intensity and task difficulty on human cognitive processing[J]. International Journal of Psychophysiology,2007,65(2):114-121 doi: 10.1016/j.ijpsycho.2007.04.001
[74] DEHAENE S,CHANGEUX J P. Experimental and theoretical approaches to conscious processing[J]. Neuron,2011,70(2):200-227 doi: 10.1016/j.neuron.2011.03.018
[75] HILLYARD S A,HINK R F,SCHWENT V L,et al. Electrical signs of selective attention in the human brain[J]. Science,1973,182(4108):177-180 doi: 10.1126/science.182.4108.177
[76] SHIFFRIN R M,SCHNEIDER W. Automatic and controlled processing revisited[J]. Psychological Review,1984,91(2):269-276 doi: 10.1037/0033-295X.91.2.269
[77] 郭志平,李安民,王积福. 乒乓球运动员的大脑功能非对称性及功能耦合性研究[J]. 天津体育学院学报,2015,30(3):256-261 doi: 10.13297/j.cnki.issn1005-0000.2015.03.014 [78] LOFFING F H N, LOFFING F, HAGEMANN N, et al. Laterality in sports: Theories and applications[M]. Amsterdam: Academic Press, 2016: 1-7
-
期刊类型引用(0)
其他类型引用(1)
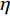
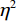


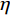
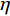
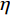

计量
- 文章访问数: 223
- HTML全文浏览量: 110
- PDF下载量: 48
- 被引次数: 1