

Precision Intervention of Targeted Neuropsychiatric Disorders Treatment: The Tapping, Application and Transformation of Exerkins
-
摘要:
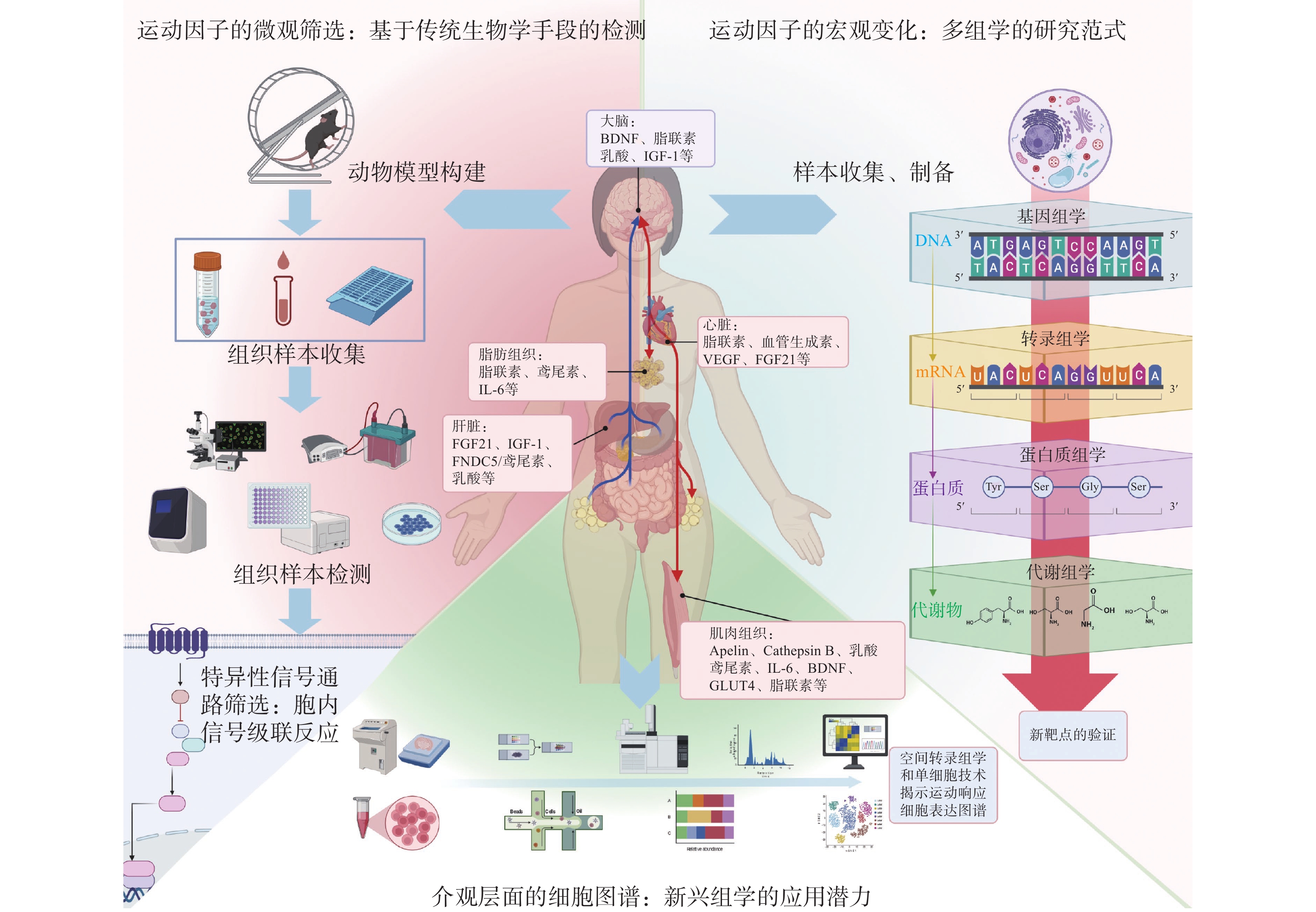
运动因子(Exerkines)作为由骨骼肌及其他组织分泌的生物活性物质,在调节神经系统功能、改善神经炎症、促进神经修复等方面具有重要作用。系统探讨运动因子的来源、转运机制及其在神经精神疾病中的作用,详细阐述运动因子的神经生物功能,并探讨其在神经精神疾病精准干预中的潜力。运动因子可分为骨骼肌源性因子、心源性因子、肝源性因子、脂肪源性因子和神经源性因子等,其神经生物学功能包括神经修复与再生、神经炎症调节、神经发育等。在运动的诱导下,这些因子通过血液循环或囊泡转运等方式传递信息,并在靶组织中发挥作用。在神经精神疾病的精准干预中,可通过运动因子进行个性化评估、精准诊断和治疗监测。通过整合生物组学、神经影像学及数据分析技术,可以更全面地了解运动因子的功能机制,并为疾病的精准干预提供科学依据。
Abstract:Exerkines, which serves as bioactive substances secreted by skeletal muscles and other tissues, play a crucial role in regulating nervous system function, improving neuroinflammation and promoting neurorepair. This study systematically explores the sources, transport mechanisms and the roles of exerkines in neuropsychiatric disorders, elaborates their neurobiological functions and the potential in precise interventions for neuropsychiatric disease treatment. Exerkines can be categorized into myokines, cardiokines, hepatokines, adipokines and neurokines. Their neurobiological functions include neural repair and regeneration, regulation of neuroinflammation and neurodevelopment. The factors induced by exercises transmit messages through blood circulation or vesicular transport, and exert effects on target tissues. In the precise intervention of neuropsychiatric disorders, exerkines can be utilized for personalized assessment, precise diagnosis and treatment monitoring. By integrating omics, neuroimaging and data analysis technologies, a more comprehensive understanding of the functional mechanisms of exerkines can be obtained, providing scientific evidence for precise intervention in diseases.
-
Keywords:
- exerkines /
- precision intervention /
- neuropsychiatric disorder /
- neurobiology
-
体育的社会分层研究伴随着现代体育发展而出现,自20世纪50年代国外学者将体育引入社会阶层理论开始,它便成为社会学领域的一个重要分支,并逐渐形成炫耀理论、社会区隔理论、社会流动理论以及冲突理论等[1−2]。随着“体育强国”“全民健身”“健康中国”等战略的出台[3],中国体育事业逐渐进入快速发展时期。在这一发展历程中,对不同社会阶层居民体育参与的研究开始进入系统研究阶段,随着人们对体育参与重视程度的提高,研究的视角也从最初的社会经济地位发展至社会空间[4]、社会资本[5]及社会网络[6]等维度,从而使研究方向呈现多元化特点。
在我国从计划经济向市场经济转型的过程中,职业是社会分层的重要基础,正如韦伯所指出的,职业是最主要且客观的阶层分化标志,不同职业群体之间具有政治观念、身份认同、经济收入、生活面向以及消费习惯等方面的边界感。因此,在当今社会发展中,它既是连接教育与收入的桥梁,又是社会声望、社会地位以及社会权利的重要来源[7]。从既有研究资料看,关于职业与体育参与关系的研究主要集中于以下3个取向:①不同职业群体的体育参与分布特征研究,如英国、荷兰以及中国经常参加体育锻炼的职业女性比例分别为44%、50%、18.43%,其中政府人员、学生等群体高于平均水平,而医务人员、企业员工、私营业主等群体低于平均水平[8]。②职业对体育参与的影响机制研究,如多数研究证实了职业地位对居民体育参与的积极影响[9],并从效能[10]、态度[11]、生涯[12]等不同机制进行了探讨。③影响职业与体育参与关系的外部因素研究,如二者的关系受到制度、市场、社会以及文化等一系列限制因素的影响[13−14]。由此来看,针对职业与体育参与关系的研究已逐渐展开,现有研究成果为深入认识职业分化对居民体育参与的影响奠定了基础。
伴随着改革开放与经济社会转型,我国经历了一个持续的职业分化过程[15]。在中国特色社会主义市场经济体制下,通过政策牵引和计划安排、劳动力市场引导与分流以及教育体系的完善,人们的就业方式以及工作单位类型呈现多样化态势,例如农民逐渐分化为雇主阶层、自雇阶层以及工薪阶层等不同职业群体[16]。在这一职业分化过程中,学者们逐渐对职业分化的概念进行了明确:随着产业结构的升级调整、社会分工的发展,人们所从事的生产活动逐渐分离成越来越细的专门性种类和专业性操作,所从事的工种日益多样化[17],如中国综合社会调查(CGSS)中所设计的党政机关、企业、事业单位、社会团体、无单位/自雇、军队等类型。
无论人们隶属于何种职业群体,均面临着2种抉择:①提高工作强度,努力赚钱;②降低工作强度,保持健康。人们参与工作的重要目的之一便是“赚钱”,这虽然能够提高人们参与体育运动的经济基础,但也会减少参与体育运动的闲暇时间,如山口周[18]指出:稍微减少劳动时间,让人们有时间享受运动流汗、音乐以及与家人聊天等,从而实现工作与生活的平衡才是建设理想高原社会所追求的。这种不同职业群体工作与生活的平衡代表了努力赚钱与闲暇时间的竞争性选择,它们之间的竞争性关系最终可能导致收入在职业分化与体育参与的关系探讨中存在非线性影响。同时,“健康”作为健康中国、全民健身等新时代背景下的“主流文化主题”,已成为不同职业群体不可或缺的目标,而居民主观健康意识的提高源于健康中国战略对主动健康的强调与宣传,无论是追求“改善健康”还是“保持健康”的行为,都体现了“主观健康”意识可能导致职业分化与体育参与产生非线性关系。
基于此,考虑到以往研究侧重探讨职业与居民体育参与的线性关系,而忽略不同职业群体对经济收入的追寻是否存在最优区间、对主观健康的认识是否在达到某个阈值时便会使职业分化对居民体育参与的影响发生改变,本文试图进一步明确以下问题:①职业分化对居民体育参与是否存在影响,这种影响在历史变迁中是增强还是弱化;②经济收入、主观健康在职业分化对居民体育参与效应中具有怎样的调节作用;③经济收入、主观健康在职业分化影响居民体育参与的过程中是否存在某个阈值或最优区间,即门限效应。在深刻理解中国的社会职业分化与体育参与关系问题的同时,根据不同职业群体的经济收入和健康追求所形成的体育参与异质性态势,提出合理政策与差异化策略,助推全民健身计划和健康中国建设发展。
1. 理论分析与研究假设
1.1 职业分化与体育参与的理论关系
健康社会学理论认为,社会成员的个体行为不仅受自身认知与态度的影响,还受宏观社会结构的限制[2]。从这一视角出发,许多学者对基于职业地位划分的社会分层与体育行为的关系进行了研究[19],其中以韦伯(经济、社会以及政治)、涂尔干(功能和职业共同体)及布尔迪厄(物质、文化以及符号)为代表所形成的社会阶层理论占据着该领域的主导地位,并衍生出各类流派。其中:炫耀理论的观点指出,体育参与群体需要有充足的时间和金钱地位,通过体育参与的某些具体项目展示和炫耀社会实力;西美尔[20]更是提出,时尚具有等级性,乃阶级分野之产物,于是体育被认为是不同职业群体社会声誉的体现。社会区隔论则强调,体育参与并不因炫耀而产生分化,而是因自身与其他职业阶层的区隔而导致居民生活的社会场域、社会空间的不同才形成体育参与分化[21]。当然,这一理论也受到一些质疑,如冲突理论认为,体育参与是一种价值观的达成,而不是阶层的区隔,体育参与的职业分化主要是因不同职业之间价值观的对立,并演化为具体体育参与上的冲突[22]。
但值得肯定的是,以上理论均认可职业分化对体育参与具有重要影响。对于高层职业群体来说,充足的时间、良好的居住环境以及向上的社交和价值观可能更有利于他们进行体育参与;而对低层职业群体来说,空闲时间有限、工作环境恶劣、居住条件差、社交匮乏以及价值观受限可能导致他们难以积极参与体育运动。当然,随着时代的变化,个体化理论指出,人们对韦伯意义上的“社会阶层”依附正在逐渐减弱[23],因为社会政策的干预、社会服务的完善以及互联网的普及等在一定程度上影响并督促着人们选择健康的生活方式(包括不饮酒、不吸烟以及体育锻炼等),如洪岩璧等[24]的研究证实了随着社会的发展以及相关政策的出台,人们在2012—2014年的饮酒等不健康行为显著下降。因此,随着时间的推移,人们将采取更加健康的生活方式来保持、改善或提高他们的健康水平,从而转向了贝克[23]所倡导的新的现代性社会发展之路,即随着我国居民生活水平的提高、居住环境的改善以及价值观的提升,职业分化对居民体育参与的影响可能在逐渐弱化。基于此,提出如下假设:
H1:职业分化对居民体育参与具有正向影响,但这种影响随着时间的变迁在减弱。
1.2 经济收入与主观健康的竞争性调节
人们在对“努力赚钱”和“主动健康”的权衡过程中可能会做出不同的体育参与行为选择,这也使在居民的体育参与系列研究中,个体经济收入与主观健康成为重要的影响变量。根据原因理论的观点,社会经济地位是造成健康行为的重要原因,收入的差异与体育参与之间存在着极大的相关性[25]。随着居民经济收入的提高,居民的消费结构将逐渐向享受型和发展型消费转变,其中体育参与(体育观赏、体育锻炼)作为重要的享受型与发展型消费行为需要良好的经济基础。Thibaut等[26]、Tovar-García[27]均验证了收入对体育参与的时间和体育消费的支出具有极强的正向影响。究其原因:①经济收入可以对人们的各类体育活动起到物质保障作用,许多体育项目均需要人们具有一定的经济条件;②经济收入达到一定水平时,人的精神和物质消费意识将被唤醒,即当人们的经济收入在承担房贷还款、教育投入、生活消费比较充足时,他们的体育消费意识才逐渐被激活。因此,居民的经济收入水平在很大程度上在职业分化与体育参与的关系中具有重要的正向调节效应。
与此同时,西方一些理论家讨论了在后现代社会中“健康”对人们的同一性建构意义,认为健康已成为一个“主流文化主题”[28]。在当前安全感相对较低的社会环境下,人们通过重视生活方式和自我提升的实践,让身体和心理成为“控制场所”,从而得到一些微小的控制感和安全感。“健康”因此成为人们感知到的现实生活中不可缺少的一种目标,而为了追寻这一目标,人们能够感知到自身健康状况的这种“主观健康”状态通常会影响其健康行为(包括体育参与、不吸烟、不喝酒等)。相关研究显示,人们的主观健康水平对体育参与具有重要影响,如王立杰等[29]认为,树立主观健康意识、进行主动健康干预是影响人们持续性体育参与的重要机制。
结合之前所论述的职业分化对居民体育参与影响的现实背景,不难发现:相比于低层职业群体的体育参与受主观健康意识影响较小,高主观健康意识的高层职业群体会比低主观健康意识的高层职业群体更容易进行体育参与[30],即便就某一职业来说,具有更高主观健康意识的群体也应该具有更好的体育参与习惯[31]。由此来看,不同职业群体对自身健康威胁的认识和对体育参与降低威胁的信念将督促他们进一步形成体育参与行为,但从既有研究资料看,对主观健康如何影响职业分化与居民体育参与的关系研究并不多见。
我国实施全民健身计划与健康中国战略以来,国家投入大量公共体育资源,从而降低底层职业群体参与体育运动的经济成本,使他们能够享受到免费、廉价的各类体育服务。因此,在健康中国战略所倡导的“主动健康”与全民健身计划所倡导的“全民健康”双重背景下,随着时间的变迁,居民体育参与的“经济成本”在逐渐降低而“健康意识”在显著提高这一事实,提出如下假设:
H2:职业分化对居民体育参与的影响因经济收入的正向调节而增强,但这种影响随着时间的变迁在减弱。
H3:职业分化对居民体育参与的影响因主观健康的正向调节而增强,且这种影响随着时间的推移更加显著。
1.3 经济收入与主观健康的门限效应
虽然以上分析探讨了个体经济收入、主观健康在职业如何影响居民体育参与中的调节效应,但是缺少分析“个人经济收入达到什么阈值才能唤醒不同职业群体的体育参与意识,主观健康状况达到什么程度才能激活不同职业群体的体育参与行为”,即“经济收入与主观健康在其中是否存在门限效应”,这也是以往研究较少涉及的更深层次的话题。在个人经济收入方面,于乐等[32]的研究指出,实物型、观赏型、健身型等体育消费水平随着收入的增加而提升,且当居民年收入达到10万元以上时,健身型与观赏型体育消费急剧增加。马斯洛[33]的需要层次理论认为,当人的某一级需求得到最低限度满足后,才会追求更高一级的需求。由此来看,当工作所获得的经济收入较低时,居民通常倾向于保守、忍耐,并减少体育参与行为,从而将自身有意识的需求控制在生存层次,而当工作所获得的收入达到一定水平后,人们才逐渐开始追求更高一级的需求,即享受层次。
当然,边际效用理论认为,当一种投入要素增加到一定程度后,所产生的效用会减少[34]。经济收入这一要素的提高对体育参与的影响也应符合这一规律。当高收入群体在追求“努力赚钱”时,忙碌的工作也会挤压他们体育锻炼的时间,从而表现出尽管某职业群体相对于其他职业群体而言的经济收入大幅提高,对体育参与的影响却在减弱。Lutfey等[35]的研究指出,个体很可能为了追求收入而牺牲体育参与等健康行为的时间,反而那些收入既达到了一定水平又不一定很高的群体,更容易腾出时间进行体育锻炼。但从既有研究资料看,很少有针对个体经济收入达到多少或者在什么门限范围内才能使职业影响居民体育参与的作用得到强化的文献。
在主观健康方面也是如此,尽管多数研究认为,人们会为了改善自己的健康状况(改善健康)或降低自身健康威胁(保持健康)而让自身的身体和心理成为在繁杂社会中能够通过主观能动性来改善的“控制场所”,但在面对“为了每月300元或500元的全勤奖仍带病坚持上班”的社会现象时,学界不得不追问:不同职业群体的这种主观健康意识达到什么程度才能突破“金钱束缚”,从而提高职业对居民体育参与的影响。由此,无论是个体经济收入还是主观健康,均可能存在门限效应。基于此,提出如下假设:
H4:个体经济收入在职业分化对居民体育参与的影响过程中存在门限效应,即当个体经济收入达到某一特定范围时,职业分化对居民体育参与的影响愈发强烈。
H5:主观健康在职业分化对居民体育参与的影响过程中存在门限效应,即当主观健康处于某一特定范围时,职业分化对居民体育参与的影响愈发强烈。
2. 研究设计
2.1 数据说明
本文的数据来源于CGSS。选取2013—2021年的数据主要是因为2013年以前的数据存在较大的结构性变化,无法形成连续性数据观察和结构分析。采用多阶段抽样,样本包括了29个省份,删除缺失值及“不知道”“不回答”等无效数据后,结合以往研究[36]对职业的划分,最终纳入无单位/自雇(包括个体户)、社会团体、企业、党政机关、军队、事业单位等6个职业群体的个案样本量为
19690 。由于本文需要分析随着时间变迁职业分化对体育参与的影响,同时考虑到自然年变化的连续属性以及后续方便对分析模型的简化与解读,将2013年赋值为1、2015年赋值为3、2017年赋值为5、2018年赋值为6、2021年赋值为9,表达实际变化年份数值,按连续变量进行处理。具体相关变量信息见表1。表 1 变量基本信息一览Table 1. List of basic variable information变量 变量类型 变量名称 统计量(编码) 2013年 2015年 2017年 2018年 2021年 因变量 连续变量 体育参与(得分) 均值 3.812 3.701 3.715 3.380 3.791 标准差 1.861 1.944 1.962 1.765 1.823 自变量 定类变量 职业群体(%) 无单位/自雇(1) 38.720 41.181 39.742 36.620 35.041 社会团体 1.310 1.392 1.091 2.650 2.302 企业 40.201 40.231 41.240 44.331 44.750 事业单位 14.475 13.310 13.481 12.405 14.130 军队 0.152 0.166 0.135 0.202 0.192 党政机关 5.142 3.720 4.311 3.792 3.585 调节变量 连续变量 个人经济收入(对数) 均值 10.425 10.012 9.553 9.734 9.876 标准差 2.654 2.420 1.145 1.135 1.215 连续变量 主观健康(得分) 均值 3.673 2.855 3.596 3.767 3.661 标准差 1.147 1.182 1.114 1.103 1.092 控制变量 分类变量 性别(%) 男(1) 48.232 45.713 51.214 50.368 46.859 女 51.768 54.287 48.786 49.632 53.141 连续变量 年龄(岁) 均值 47.385 48.208 48.982 48.657 50.503 标准差 15.703 16.119 16.075 16.256 16.783 分类变量 婚姻(%) 未婚(1) 9.684 9.597 8.806 10.268 10.344 已婚 90.316 90.403 91.194 89.732 89.656 分类变量 户口状况(%) 农业(1) 51.409 56.396 53.303 55.436 56.752 非农业 48.591 43.604 46.697 44.564 43.248 连续变量 受教育程度(等级) 均值 4.955 4.767 4.935 4.978 4.942 标准差 2.856 2.943 3.117 3.139 3.093 (1)因变量。体育参与是指人们为了更好地实现身心健康、活跃文化生活、加强社会交往,采用体育锻炼、娱乐休闲等方法进行的有意识、有计划的体育行为,包括直接参与和间接参与[37]。选取具有直接参与性的“体育锻炼”和间接参与性的“观看比赛”2个指标构成体育参与行为。根据CGSS问卷数据库中的题项,“过去一年,您是否经常在空闲时间从事以下活动?”其中包括“参加体育锻炼”和“观看体育比赛”2个选项,回答有“每天”“一周数次”“一月数次”“一年数次或更少”“从不”5个选项,对应的编码是1~5。统计时为了使编码与体育参与程度的方向一致,采用y=−x+6的方式对原始数据进行变量转换,同时,将“参加体育锻炼”和“观看体育比赛”2个变量的选项进行合并编码,从直接参与和间接参与2个方面形成本文中体育参与的变量值,按连续变量处理,运用OLS回归分析。
(2)自变量。职业是指个人所从事的服务于社会并作为主要生活来源的工作[15]。CGSS数据调查中将职业划分为无单位/自雇(包括个体户)、社会团体、企业、事业单位、军队、党政机关和其他,统计分析中未考虑“其他”类别,将以上6种职业按1~6进行编码统计,属于定类变量。
(3)调节变量。①个人经济收入(年收入)。CGSS调查中的题项为“您个人去年全年的职业/劳动收入是多少?”主要反映社会经济地位,在统计中采用自然对数进行数据转换,属于连续变量。②主观健康。CGSS调查中的题项为“您觉得您目前的身体健康状况是?”回答从“很不健康”到“很健康”分别赋值1~5分。美国医学社会学相关论文[38]大多有将自评健康采用1~4编码时处理为连续变量的传统,本文延续这一规范,也将其处理为连续变量。
(4)控制变量。主要包括5个变量:①性别,统计中男性编码为1、女性编码为2;②年龄;③婚姻,将调查中的未婚设为1,初婚有配偶、再婚有配偶、分居未离婚、离婚、丧偶等编码为2;④户籍状况,设农业户口为1,非农业户口、蓝印户口、居民户口、军籍等为2,剔除没有户口和其他选项;⑤受教育程度,删除“无法回答”和“其他”选项后,将未受过任何教育、私塾、小学、初中、职业高中、普通高中、中专、技校、大学专科(成人高等教育)、大学专科(普通高等教育)、大学本科(成人高等教育)、大学本科(普通高等教育)、研究生及以上分别编码为1~13。
2.2 模型设置
模型一:时间固定效应模型(公式1)。以体育参与为因变量,以职业为自变量,以发展年份为时序变量,在控制影响体育参与的其他因素基础上构建多元回归模型。在按年代时序验证职业对体育参与影响变化的同时,为了估计体育参与随时间变迁的轨迹和因个人经济收入、主观健康导致的体育参与轨迹异质性,采用如下回归模型:
$$ \begin{split} {\mathrm{SP}}_{it}=&{\mathrm{\beta }}_{00}+{\mathrm{\beta }}_{01}{\mathrm{career}}_{it}+{\mathrm{\beta }}_{20}{\mathrm{income}}_{it}+{\mathrm{\beta }}_{02}{\mathrm{health}}_{it}+\\&{\mathrm{\beta }}_{21}{\mathrm{career}}_{it}\times {\mathrm{income}}_{it}+{\mathrm{\beta }}_{03}{\mathrm{career}}_{it} \times {\mathrm{health}}_{it}+\\&{\mathrm{\beta }}_{10}{\mathrm{year}}_{it}+{\mathrm{\beta }}_{11}{\mathrm{career}}_{it}\times {\mathrm{year}}_{it}+{\mathrm{\beta }}_{21}{\mathrm{income}}_{it}\times {\mathrm{year}}_{it}+\\&{\mathrm{\beta }}_{30}{\mathrm{health}}_{it} \times {\mathrm{year}}_{it}+{\mathrm{\beta }}_{13}{\mathrm{career}}_{it}\times {\mathrm{income}}_{it}\times {\mathrm{year}}_{it}+\\&{\mathrm{\beta }}_{31}{\mathrm{career}}_{it}\times {\mathrm{health}}_{it}\times {\mathrm{year}}_{it} +\sum _{j > 4}{\mathrm{\pi }}_{ji}({{{X}}_{{j}})}_{it}+\\&\sum _{j > 1}{\mathrm{\beta }}_{0j}({{{Z}}_{j})}_{i}+{{e}}_{it}\\[-1pt] \end{split} $$ (1) 通过Hausman检验(Chi2=605.61,P<0.05),选择固定效应模型(表2)。SPit(sports participation)是衡量体育参与的指标,代表不同年份的体育参与情况;yearit是体育参与的不同年份,由于此处年份已作为数值性数据处理,所以统计过程中已进行中心化处理(减去平均年份);careerit、incomeit、healthit分别代表不同职业、收入水平、主观健康指标,其中个人经济收入用对数表示;β代表模型参数,πji代表随时间变化的控制变量的模型参数,
$ {{X}}_{j} $ 是随时间变化的控制变量,$ {{Z}}_{j} $ 是不随时间变化的控制变量,$ {{e}}_{it} $ 是个体i在时间t的残差。同时,考虑到回归模型分析中的异方差问题,具体统计过程中使用稳健标准误进行回归分析(robust命令)。表 2 Hausman检验结果Table 2. Hausman test results变量 (b)
固定效应(fe)(B)
随机效应(re)(b-B)
Differencesqrt(diag
(V_b-V_B))
S.E.职业2 0.303 0.342 −0.039 0.005 职业3 0.195 0.261 −0.066 0.010 职业4 0.392 0.430 −0.038 0.012 职业5 1.169 1.223 −0.054 0.014 职业6 0.460 0.476 −0.016 0.009 个人经济收入 0.086 0.115 −0.029 0.002 主观健康 0.075 0.072 0.003 0.002 注:职业1表示无单位/自雇职业群体,为参照组;各效应的控制变量均已得到控制;Chi2为卡方检验的统计量,Prob > Chi2表示在给定的显著性水平下,观察到的卡方值或更大的值出现的概率。如果Prob > Chi2的值小于显著性水平(通常是0.05),则拒绝原假设。本文Prob > Chi2的值小于 0.0001 ,因此模型具有显著性。模型二:门限模型(公式2和公式3)。门限回归模型旨在处理解释变量与被解释变量的非线性关系和结构突变问题,它通过设定一个或多个门限变量及对应的阈值,将数据分割成不同区间,对每个区间内的数据进行线性回归分析,从而使得整个模型能够有效地描述变量之间的非线性关系,这一模型目前已广泛应用于经济学领域[39]。本文需要分析个人经济收入与主观健康达到什么阈值才能激发不同职业群体的体育参与行为,从哪些区间划分又能够表现出它们之间的非线性关系,而这一目的的实现正是门限回归模型分析方法的优势所在,通过这一模型深入挖掘个体经济收入和主观健康在不同状态区间的预估方程,有利于突破以往单纯以线性回归造成伪回归效应的局限。具体方程模型(在Stata分析中以3个门限值为先的原则进行逐次递减建模)如下:
$$ {Y_{it}} = {\alpha _i} + {Z_{it}}\delta + 1(q{}_{it} \leqslant \gamma )X_{it}^{'}{\beta _1} + 1({q_{it}} > \gamma )X_{it}^{'}{\beta _2} + {\varepsilon _{it}} $$ (2) 其中,q代表门限变量,γ代表具体门限值,ε代表随机扰动项,δ代表不随时间变化的控制变量参数,δ不受门限值的影响,
$X' $ 代表随时间变化的控制变量,示性函数$ 1( \cdot ) $ 。$$ 1({q_{it}} \leqslant \gamma ) = \left\{ \begin{gathered} 1\quad{\text{ if }}{q_{it}} \leqslant \gamma \\ 0\quad{\text{ if }}{q_{it}} > \gamma \\ \end{gathered} \right.;{\text{ }}1({q_{it}} > \gamma ) = \left\{ \begin{gathered} 0\quad{\text{ if }}{q_{it}} \leqslant \gamma \\ 1\quad{\text{ if }}{q_{it}} > \gamma \\ \end{gathered} \right. $$ (3) 3. 研究结果
3.1 主效应与调节效应检验
进行回归分析时选取自变量容差和反差膨胀因子(VIF)对多重共线性进行检验,结果发现各指标VIF小于2,各容差大于0.1,表明自变量均满足要求,不存在多重共线性问题。
表3中的模型1显示,职业对体育参与产生了显著的积极影响,相比于无单位/自雇群体,工作于企业的群体提高体育参与0.273个单位,服务于社会团体的群体提高体育参与0.392个单位,就职于事业单位和党政机关的群体分别提高体育参与0.488和0.462个单位,明显高于企业的影响程度,而军队群体由于自身职业属性,提高体育参与达1.273个单位,以上结果验证了体育参与的职业分化现象。同时还发现,职业对体育参与的影响随着年度的变化呈显著的负向变化(
$ {\beta }_{11} $ =−0.026,P<0.01),表明职业分化对体育参与的影响作用正在减弱,说明尽管军队、党政机关、事业单位等职业群体可能比无单位、自雇阶层等职业群体具有更良好的工作环境、更多的社会资源和闲暇时间,但随着我国全民健身计划的进一步实施,亲民性、大众化以及生活化的公共体育服务政策与设施逐渐完善,空间的标准化、精准化、均等化等结构供给与调整逐渐优化,这一职业分化所带来的差异正在缩减。H1得到验证。表 3 职业分化对体育参与影响的效应模型Table 3. The effect model of occupational differentiation on sports participation固定效应 模型1 模型2 模型3 模型4 模型5 截距模型 截距 2.741***
(0.071)1.402***
(0.150)2.512***
(0.091)1.159***
(0.179)1.079***
(0.162)职业(无单位/自雇=1) 社会团体 0.392***
(0.110)0.388***
(0.102)0.379***
(0.103)0.412***
(0.128)0.468***
(0.141)企业 0.273***
(0.301)0.272***
(0.047)0.269***
(0.030)0.328*
(0.155)0.375*
(0.161)事业单位 0.488***
(0.054)0.506***
(0.048)0.455***
(0.043)0.590*
(0.248)0.659**
(0.231)军队 1.273***
(0.301)1.175***
(0.291)1.231***
(0.279)1.289**
(0.446)1.421**
(0.458)党政机关 0.462***
(0.057)0.543***
(0.081)0.486*
(0.059)0.681*
(0.417)0.845*
(0.421)个人经济收入(对数) 0.132***
(0.014)0.130***
(0.018)0.139***
(0.020)主观健康 0.089***
(0.015)0.082**
(0.023)0.075*
(0.021)职业×收入 0.079**
(0.006)0.075*
(0.007)职业×健康 0.066*
(0.013)0.060*
(0.010)线性增长模型 年份 −0.018
(0.013)−0.301***
(0.057)−0.015
(0.005)−0.260***
(0.071)−0.280***
(0.075)职业×年份 −0.026**
(0.004)−0.024**
(0.005)−0.022**
(0.005)−0.021*
(0.005)−0.020*
(0.005)收入×年份 0.026***
(0.007)0.028***
(0.008)0.037***
(0.009)健康×年份 −0.015
(0.006)−0.014
(0.008)−0.013
(0.009)职业×收入×年份 −0.018*
(0.003)职业×健康×年份 0.006
(0.003)R2 0.1528 0.1553 0.1529 0.1562 0.1571 F 260.22** 203.14** 200.58** 172.15** 150.49** 注:*表示P<0.05,**表示P<0.01,***表示P<0.001;括号内为稳健标准误;各模型中的控制变量均已经得到控制。 就个人经济收入和主观健康而言,两者均对居民的体育参与行为具有积极的正向影响,且从模型2和模型3的对比来看,个体的主观健康意识每提高1个单位,居民的体育参与程度提升0.089个单位,而代表经济水平的个人收入(
$ {\beta }_{20} $ =0.132,P<0.001)对居民体育参与的影响要显著高于主观健康因素。从个体经济收入、主观健康与职业的交互效应看,具有高收入的职业群体更容易形成体育参与习惯($ \beta_{21} $ =0.079,P<0.01),具有高主观健康评价与意识的职业群体也更容易形成体育参与习惯($ {\beta }_{03} $ =0.066,P<0.05),但个体经济收入比主观健康在职业与体育参与的关系中具有更强烈的影响效果。进一步进行三阶调节效应检验,分别将职业、个体经济收入以及年份的乘积,职业、主观健康以及年份的乘积加入回归,发现前者对体育参与的影响仍然显著($ {\beta }_{13} $ =−0.018,P<0.05),而后者对体育参与的影响不显著($ {\beta }_{31} $ =0.006,P>0.05)。以上结果说明,尽管收入越高的职业群体更容易形成体育参与习惯,但这种效应正在随着时间的变迁而弱化,而主观健康在其中的积极影响却不随时间的变迁而改变。因此,研究结果验证了H2,推翻了H3。3.2 门限效应检验
为了避免上述线性模型造成的伪回归现象,需要对本文中涉及的个人经济收入和主观健康进行门限效应分析,验证是否存在门限值,然后根据门限值范围进行分段回归或预估,才能正确了解在职业分化影响居民体育参与中“经济收入”和“主观健康”起到的深层作用。根据三门限效应验证原理进行门限效应检验,结果(表4)显示:主观健康存在单门限效应(F=129.150,P<0.001),双门限效应不显著(F=66.892,P>0.05);经济收入存在单门限效应(F=523.110,P<0.001)和双门限效应(F=168.052,P<0.001),三门限效应不显著(F=16.385,P>0.05)。以上结果通过了构造似然比函数和95%置信区间的检验。为此,需要根据门限估计值分别进行分段预估(主观健康需要分析γ≤3.000和γ>3.000这2个阶段在职业对居民体育参与影响中的异质性;经济收入需要分析γ≤8.847、8.847<γ≤9.962以及γ>9.962这3个阶段在职业对居民体育参与影响中的异质性),以避免对数据的绝对均衡化要求。
表 4 门限效应检验Table 4. Threshold effect test模型 门槛估计值(γ) F P BS 临界值 95%置信区间 1% 5% 10% 主观健康 单门限 3.000 129.150 <0.001 300 15.175 8.316 6.709 [2.100,4.000] 双门限 4.000 66.892 1.000 300 9.597 6.127 4.834 [3.500,5.000] 经济收入对数 单门限 9.962 523.110 <0.001 300 14.957 10.828 8.919 [9.814,9.985] 双门限 8.847 168.052 <0.001 300 15.198 10.588 9.069 [8.736,8.896] 三门限 9.298 16.385 0.225 300 60.265 38.187 21.852 [9.256,9.314] 分段预估结果如表5所示。就主观健康门限(γ≤3.000和γ>3.000)来看,无论处于何种门限范围,相较于无单位/自雇群体,其他职业对居民体育参与均表现出显著影响。但令人意外的是,当人们的主观健康状况评价在γ≤3.000范围时,与γ>3.000范围比较,除军队以外的其他职业均表现出对居民体育参与的影响更强。进一步将职业、健康结合年份进行交互效应分析发现,随着时间的变迁,主观健康的调节效应并未增强(
$ {\mathrm{\beta }}_{31} $ =−0.039,P>0.05)。由此看来,主观健康在职业分化与体育参与的关系中起到了二阶段调节效应,第一阶段(γ≤3.000)的调节效应强于第二阶段(γ>3.000),且这种影响并不随时间的变迁而变化。表 5 职业分化对体育参与影响的门限效应全模型Table 5. Full model of threshold effect of occupational status on sports participation固定效应 健康门限模型 经济收入门限模型 γ≤3.000 γ>3.000 γ≤8.847 8.847<γ≤9.962 γ>9.962 截距模型 截距 1.020***
(0.785)0.881**
(0.459)0.473*
(3.919)1.071**
(0.258)0.951**
(0.655)职业(无单位/
自雇=1)社会团体 0.570**
(0.486)0.532**
(0.279)0.061
(0.016)1.221***
(0.389)0.598**
(0.169)企业 0.658**
(0.758)0.641**
(0.456)0.070
(0.029)1.458***
(0.611)0.632**
(0.258)事业单位 1.565***
(1.157)0.780***
(0.188)0.066
(0.022)3.231***
(1.527)1.274*
(0.666)军队 0.851**
(1.963)1.614***
(1.038)0.852*
(0.048)2.605***
(1.359)1.590*
(0.648)党政机关 1.133**
(0.539)0.881**
(1.915)0.060
(0.021)2.338***
(0.919)1.026*
(0.388)个人经济收入
(对数)0.180*
(0.083)0.232***
(0.041)0.088
(0.026)0.183***
(0.027)0.315***
(0.075)主观健康 0.110**
(0.167)0.091*
(0.087)0.511*
(0.382)0.570*
(0.029)0.530*
(0.059)职业×收入 0.119**
(0.029)0.082*
(0.012)0.048
(0.100)0.104*
(0.019)0.080*
(0.011)职业×健康 0.089**
(0.028)0.060*
(0.015)0.064*
(0.017)0.067*
(0.010)0.081*
(0.016)职业×收入×年份 −0.032*
(0.014)−0.023*
(0.004)0.035*
(0.249)−0.010
(0.005)−0.028*
(0.008)职业×健康×年份 −0.039
(0.028)−0.038
(0.024)−0.058
(0.128)−0.015
(0.005)−0.030
(0.009)R2 0.1870 0.1616 0.2803 0.1605 0.1476 F 281.23** 264.25** 297.41** 231.55** 207.19** 注:*表示P<0.05,**表示P<0.01,***表示P<0.001;括号内为稳健标准误;各模型中的控制变量均已得到控制。 从经济收入门限看,当人们处于低收入(个体经济收入在第一门限以下,γ≤8.847)时期时,相比于无单位/自雇群体,除军队职业群体外,其他职业群体与其在体育参与上无显著差别。尽管此阶段收入不占主导地位(各职业群体均处于低收入阶段,难以支撑体育参与),但随着时间的变迁,收入在职业分化对体育参与的影响过程中的作用逐渐增强(
$ {\mathrm{\beta }}_{13} $ =0.035,P<0.05)。处于中收入(经济收入在第一门限到第二门限之间,8.847<γ≤9.962)时期时,居民的体育参与逐渐表现出职业分化,且随着时间的变迁,这种分化现象保持不变($ {\mathrm{\beta }}_{13} $ =−0.010,P>0.05)。处于高收入(经济收入在第二门限以上,γ>9.962)时期时,尽管职业分化也表现出对居民体育参与具有显著影响,但结果显示这种影响在逐渐减弱($ {\mathrm{\beta }}_{13} $ =−0.028,P<0.05)。这说明“努力赚钱”确实在一定程度上牺牲了体育参与等健康行为的时间,反而那些中收入职业群体更容易进行体育参与。由此来看,经济收入在职业分化与居民体育参与的关系中起到三阶段调节作用,即当居民的经济收入在8.847万元及以下时,相比于无单位/自雇群体,除军队外,其他职业对居民的体育参与不存在显著影响,但随着时间的变迁有明显的增强趋势;在8.847万元<经济收入≤9.962万元时,与无单位/自雇群体比较,各职业对居民体育参与的影响表现出显著性,且这种影响不随时间的变迁发生变化;经济收入在9.962万元以上时,职业分化仍对居民体育参与产生显著影响,但有明显的减弱趋势。
以上结果说明职业分化对居民体育参与的影响在主观健康与个体经济收入上均存在门限效应:前者呈现出单门限效应,分为健康状况一般阶段(γ≤3.000)和健康状况良好阶段(γ>3.000);后者表现出双门限效应,分为经济收入不高于8.847万元阶段、8.847万元<经济收入≤9.962万元阶段以及经济收入高于9.962万元阶段。以上结果验证了H4和H5。
4. 讨 论
职业是人们获取劳资报酬的重要手段,“努力赚钱”一直是不同职业群体养家糊口或追求更高生活品质的重要途径。在健康中国战略强调“主动健康”的背景下,“健康”已然成为社会职业群体在工作中的一项“重要控制目标”,“要职业不要职业病”似乎使“努力赚钱”与“保持健康”成为一种对立关系,但这也使得“职业与健康行为”之间的话题变得更加有趣。其实,无论是以韦伯的经济地位、政治地位以及社会荣誉三位一体形成的社会分层模式,还是以布尔迪厄物质资本、文化资本以及符号资本划分的社会分层类型,抑或是涂尔干用职业共同体来解释社会现象以及人们的社会行为,其代表的各维度社会显性结果均指向了职业分化。这是因为社会职业不仅反映了个体的受教育程度,还在一定程度上表达着个体的家庭背景、经济地位以及社会话语权,从而代表了制度、结构、思想以及行动上的差异[2]。正是因为职业在很大程度上影响了以上社会现象及社会行为,必然也影响着人们体育参与行为的发生,从而使本文的研究结果表现为职业分化对居民的体育参与具有显著性影响,且随着社会经济的不断发展,这种影响力在逐渐减弱。
究其原因:①与拉德克利夫-布朗[40]所强调的社会结构有关,该理论指出社会结构是影响并制约个体的外部环境,如国家政策。在国家政策的推动下,我国提出了全民健身计划、体育强国战略以及健康中国战略,其中全民健身计划强化了国民体质监测、体育锻炼标准以及场地设施标准等,为不同群体提供了标准化的体育服务[41];体育强国战略强调了“逐步推动基本公共体育服务在地区、城乡、行业和人群间的均等化”[42];健康中国战略在强化“完善全民健身公共服务体系”的同时,还进一步强调了“到2030年,健康生活方式得到普及······基本实现健康公平”的战略目标[43]。正是由于国家政策的推动,社会体育资源的配置逐渐延伸到不同的社区、村落等,从而提高了不同职业群体社会资源占有的公平性,弱化了经济收入在其中的关键作用,最终使职业分化对居民体育参与的影响逐渐减弱。②与拉波特等[44]提出的个体能动性有关,该理论强调人是能动性的个体,在这里则表现为人们对健康的认知以及体育行为的预期。作为具有能动性的个体,不仅认识到体育不仅能够提高生理、心理以及社会适应等层面的整体健康状态,还体会到体育也是个体从“自然人”向“社会人”转变的重要手段之一,具有为他们自身优化社会网络、扩展社会资本以及提高社会信任等功能,而这提高了各职业群体的体育参与行为,减弱了职业对体育参与的影响,最终表现为职业分化对体育参与的影响随时间的变迁而减弱。
针对核心议题,职业群体追寻的经济收入在什么区间,主观健康达到哪一阈值时,职业分化对居民体育参与的影响会发生变化,研究展示了不同的结果。就主观健康的阈值而言,结果发现主观健康存在单门限效应,表现为在职业分化与体育参与关系中具有二阶段调节作用,即在职业对体育参与的影响中,居民主观健康在第一状况阶段(γ≤3.000)比在第二状况阶段(γ>3.000)具有更高的调节效应。这是由于人们在主观健康评价为“很不健康”(γ=1)时,通常伴随着“生理疾病”的出现,此时难以进行体育参与;当人们在主观健康评价为“不太健康”(γ=2)或“一般健康”(γ=3)时,通常表现为心理健康上的“压力”“抑郁”“焦虑”“失眠”以及社会适应上的“冲突”“交往”“自控力”等方面的健康问题。此时人们更愿意选择通过体育参与行为来调整心理压力、缓解心理抑郁以及提高社会适应,而这种从“很不健康”(生理疾病的出现导致难以进行体育参与)到“一般健康”(心理释放与社会适应的需求导致愿意选择体育参与)的显著变化,必然表现出“体育参与”在其中表现出更加强烈的调节效应。同时,本文结果表明了相对于无单位/自雇、企业、社会团体等职业群体,事业单位、党政机关更容易进行体育参与行为。这些群体通常具备更高的教育水平,掌握着良好的健康保健知识与体育教育资源,因此,为了改善健康,他们更容易形成体育参与习惯。
研究还发现,在健康的第二状况阶段,相比于无单位/自雇群体,除军队职业群体外,其他职业群体尽管表现出明显的体育参与倾向,但这一关系均不如第一状况阶段影响强烈,表明此阶段职业群体的目的与第一阶段存在差异。当居民主观健康评价为“健康”(γ=4)和“很健康”(γ=5)时,居民更想通过体育参与来保持健康。当然,居民的主观健康无论处在哪一水平,均未表现出随着时间的变迁而增强的态势,说明我国居民主动健康意识仍显不足。究其原因:①与教育体系不完善有关。健康教育在学校的覆盖率和深度相对不足,由于缺乏系统、长效的健康教育,人们的主动健康意识欠缺。②与医疗体系的限制有关。中国居民通常受此限制,习惯被动接受医疗服务,而缺乏主动预防和健康管理意识。③与社会支持系统缺陷有关。目前我国缺乏有效的社会支持和健康管理社区网络,使得个体难以获取健康信息,导致健康意识提升不足。④与个体态度有关。由于不同职业群体的高强度工作压力,个体对健康的认识和关注不足,导致难以形成主动健康意识。
未来,进一步提高我国居民的主动健康意识将是减少体育参与职业分化的重要一环。由此来看,主观健康在居民职业分化与体育参与关系中的这种二阶段调节效应表现为:居民健康状况欠佳(第一阶段)时,通常想通过体育参与来改善健康;当居民健康状况良好(第二阶段)时,则想通过体育参与来保持健康。
其实,尽管当今社会职业地位存在巨大差别,但在事业单位、党政机关、企业、自雇等群体中整体收入差别不大,甚至有些职业地位尽管偏低,但收入相对较高,如有些街头小贩、外卖员的收入并不亚于企事业单位员工。因此,尽管职业与经济收入存在相关性,但其并未保持一种严格意义上的线性关系,因此有必要探讨经济收入在职业分化影响居民体育参与的过程中起到怎样的深层作用。
本文结果显示,个体经济收入在职业分化与体育参与关系中具有三阶段调节效应,即:当居民的经济收入在8.847万元及以下时,与无单位/自雇群体比较,除军队群体外,其他职业群体与其职业分化对体育参与不存在显著影响,但有明显的增强趋势;当8.847万元<经济收入≤9.962万元时,职业分化对居民体育参与表现出积极影响,且这种影响随着时间的变迁保持不变;当经济收入在9.962万元以上时,职业分化对居民体育参与的影响仍然持续,但有明显的减弱趋势。在以上变化过程中需要注意:不同职业群体的“主观健康”意识一直具有积极的调节效应,即便在低收入阶段职业分化对体育参与的无影响时期和高收入阶段职业分化对体育参与影响的弱化时期,通过提高“主观健康”意识来提升不同职业群体的“主动健康”行为亦是一种良好的选择。
以上结果可由经济学中的消费者选择理论和社会学中的贝克尔境界理论综合来解释。贝克尔境界理论指出,人们的每项任务都有一个最佳唤醒点[45]。当居民的收入水平尚未达到最佳点时,便无法唤醒他们的体育参与行为,由此表现出居民收入偏低(处于第一阶段)时,职业分化对居民体育参与的影响不显著,表现为大部分职业群体如果收入较低,其认知、行为和消费处于生存型状态,则难以选择属于享受型和发展型的体育观赏、体育锻炼等行为方式;当居民收入达到一定水平(最佳唤醒点)时,居民的体育参与水平便会明显提升。这正与消费者选择理论的观点相衔接,该理论强调消费者的收入水平对购买商品或服务有着重要影响[46],从而使居民的经济收入达到第二阶段时,个体经济收入表现出极强的调节效应。但该理论同时还指出,这种影响存在边际效用,当收入达到某个点时,由于服务水平的限制、时间成本的提高以及社会资本的分配,对购买某项产品或服务将会缩减,即当居民的经济收入处于第三阶段时,他们的经济收入水平在职业分化对居民体育参与的影响中的效应将减缓。这里需要指出的是,尽管未来不同的研究针对不同的调查样本可能得出不同的拐点,但笔者认为:学界更应该强调经济收入在职业分化与体育参与的关系中具有“三阶段调节效应”的学理性观点,以此来规划低收入职业群体的公共体育服务、优化高收入职业群体的精准性体育服务以及聚焦居民的“主动健康”意识培养等相关政策,因而丰富与完善职业分化与体育参与的理论与治理体系是关键。
5. 结论、建议与局限
5.1 结 论
本文在“努力赚钱”与“主动健康”的社会发展现象中,从个体经济收入与主观健康的视角分析了职业分化对居民体育参与的影响。结果发现:①职业分化对居民体育参与具有积极的正向影响,但这种影响随着时间的变迁正在减弱;②主观健康在职业分化与体育参与关系中存在单门限效应,表现出二阶段调节作用,分别表达了人们在第一阶段追求改善健康和在第二阶段追求保持健康的目的,但这种影响并未随着时间的变迁而得到强化;③个体经济收入在职业分化与体育参与关系中存在双门限效应,表现出三阶段调节作用,从而使体育参与的职业分化现象随着时间的变迁呈现出“增强—不变—弱化”的阶段性变化;④无论是在职业分化对居民体育参与的“无影响阶段”(低收入阶段),还是“弱化影响阶段”(高收入阶段),提高人们的主观健康意识从而促进主动健康行为都将起到重要的弥补作用。
5.2 建 议
(1)预计2025年全民健身计划将进一步提高体育锻炼人口,但职业分化在这一过程中仍然是影响人们体育参与的重要变量,说明应进一步强调不同职业群体公共体育服务的均等化发展,尤其加强对弱势职业群体的关注是提高公共体育服务均衡化发展的重要一环。因此,需要更深入地通过“人—财—物”资源整合提高服务的高效性;通过场地合理规划与构建“举步可就”的健身设施提高服务的均等化;通过交通联动优惠等全民健身政策提高服务的便捷性;通过开设特定体育课程、开展“运动银行”等策略提高服务的多元化;通过动态资源调配、智能推荐系统开发等路径提高服务的全面性。
(2)高收入职业群体由于公共体育服务水平的限制、自身时间成本的提高以及社会资本的分配,随着时间的推移趋向于缩减体育参与,因此需要进一步通过制定高品质的、舒适的个性化体育服务,时间灵活的全天候健身环境以及持续跟踪的综合性反馈机制等来提高针对高收入职业群体的精准性体育服务水平。
(3)尽管健康中国战略强调“主动健康”,但研究显示中国居民的“主动健康”意识仍显不足,因此未来应构建完善的医疗体系、教育系统以及社会支持网络,并通过赛事引领、体育网络熏染、体育自媒体宣传等手段,进一步提升全民的主动健康意识,促进居民“主动健康”行为素养的形成。
5.3 局 限
本文将经济学研究中的门限效应模型引入体育学研究,对职业分化与体育参与关系的挖掘有了新的发现,但同时也存在如下研究局限:①尽管探讨了职业分化与体育参与的直接效应、调节效应以及门限效应,但缺乏对中介效应的考量,未来可从具有职业属性的工作强度和个体属性的自我效能等中介路径进行深入探讨。②体育参与的自评式题项可能存在一定的主观性,未来如能采用客观的体育参与“频率”“时间”“强度”来对其进行综合评价将是提高研究客观性与严谨性的关键,同时这也提示着广大体育学人,体育与健康有关数据库的构建是本领域未来亟待发展的一个重要方向。
作者贡献声明:刘微娜:提出论文主题,设计论文框架,撰写、修改论文;作者贡献声明:章森、何文柯:协助检索、筛选、梳理文献,协助修改论文;作者贡献声明:漆正堂:提出论文主题,设计论文框架,审核、指导修改 论文。 -
![]()
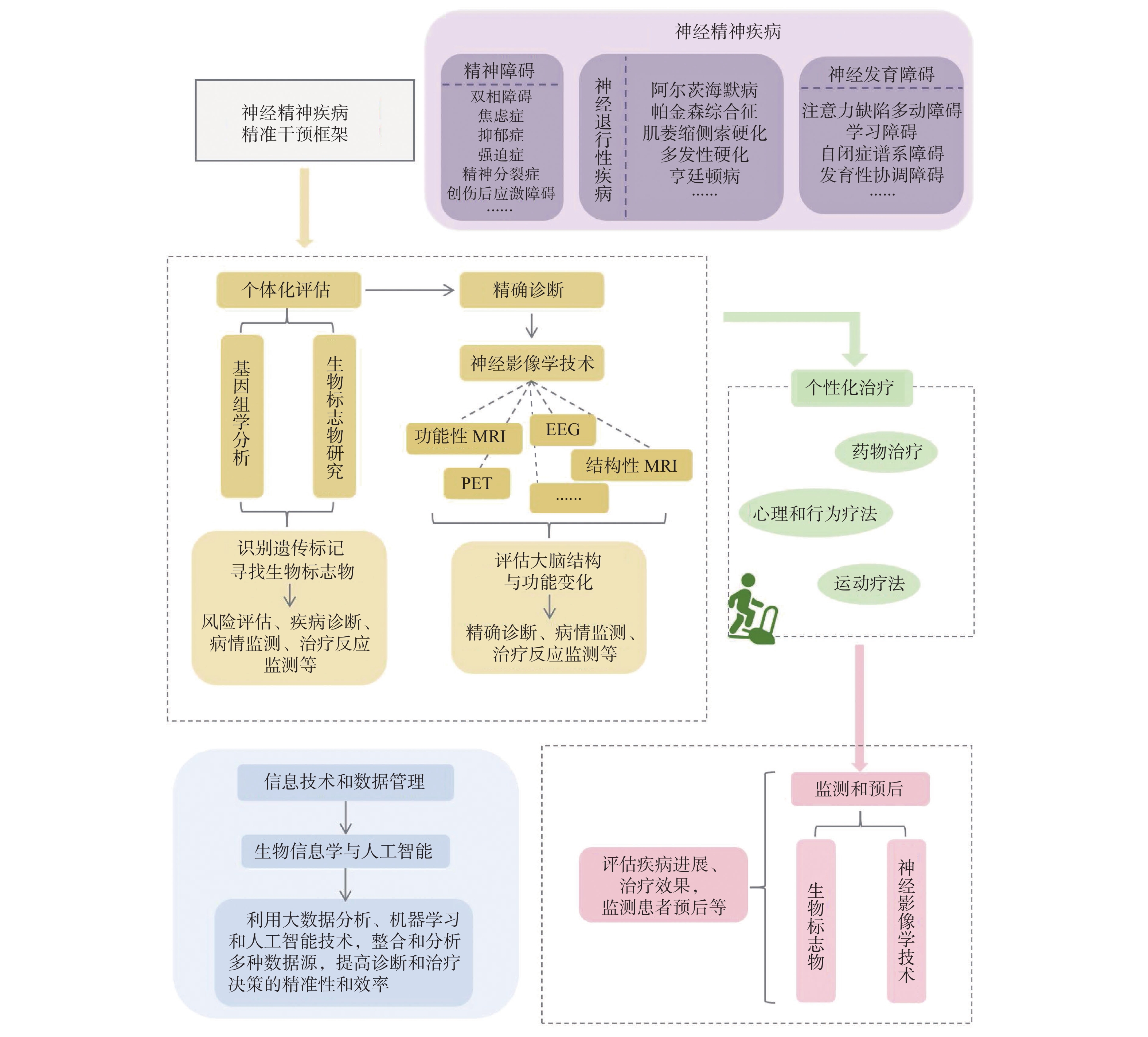
图 2 神经精神疾病的运动干预框架
Figure 2. Exercise intervention framework for neuropsychiatric disorders
![]()
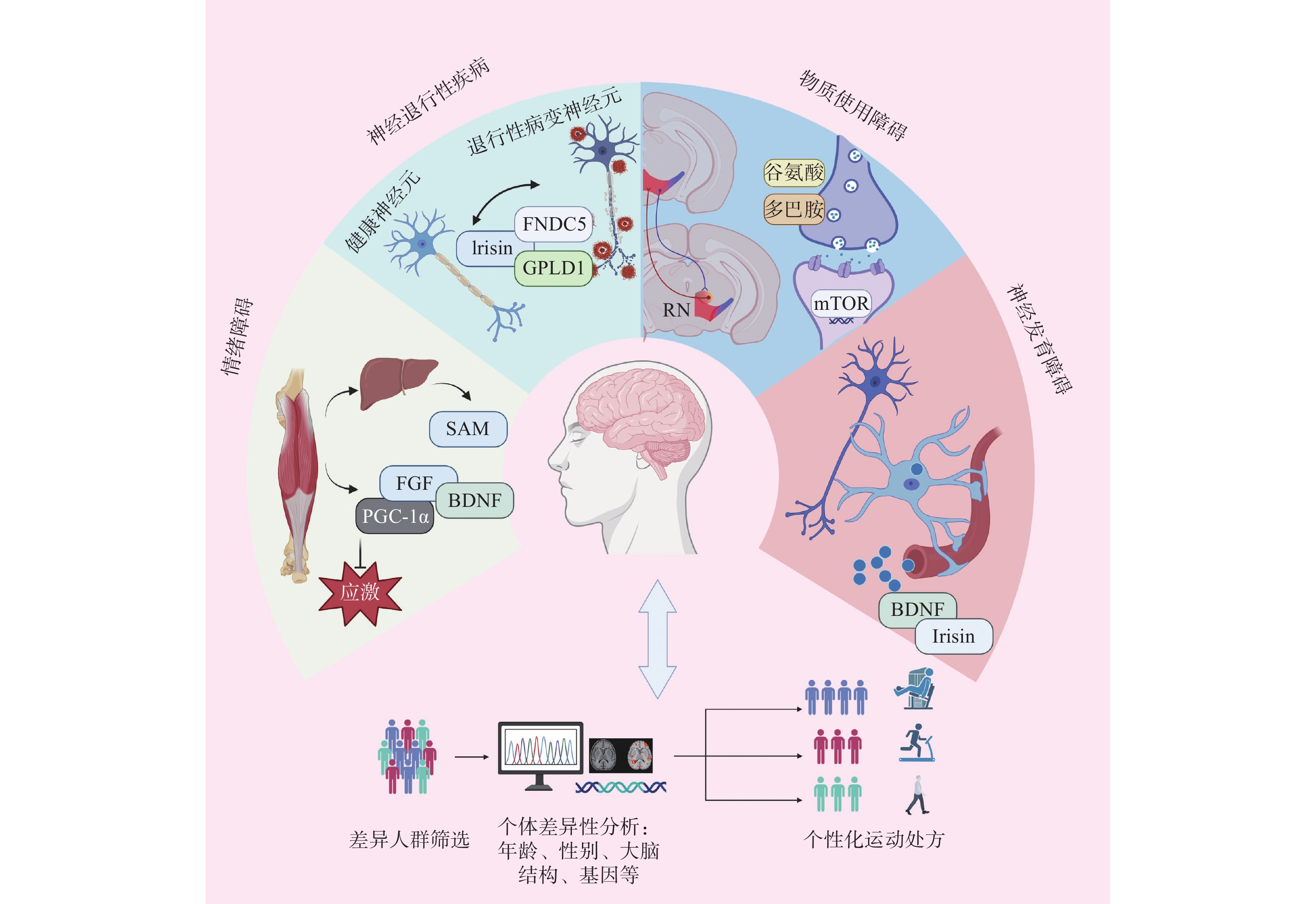
图 4 运动因子在神经精神疾病干预中的应用
Figure 4. Application of exerkines in the intervention of neuropsychiatric disorders
表 1 主要运动因子的来源、转运及靶向组织
Table 1 Sources, transport, and target tissues of major exerkines
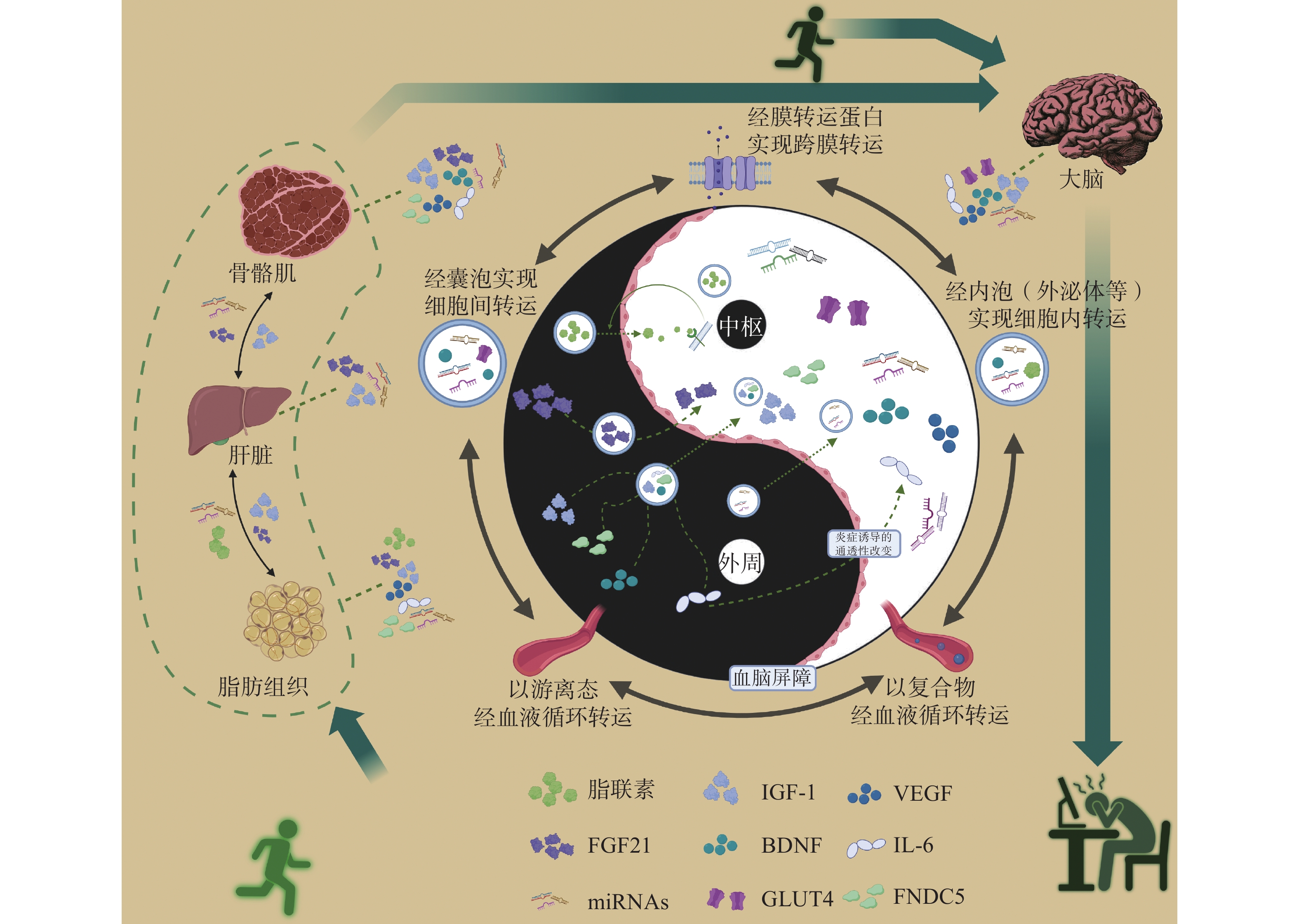
运动因子 主要来源/表达组织 转运方式/信号转导方式 神经生物学功能 脂联素 骨骼肌[16]、脑[18]、
脂肪[23]由囊泡运输至胞外[23],经血液循环运送[24],与靶细胞上的特异性受体结合进行信号转导[25] 抗氧化、抗凋亡[25]
增强突触可塑性[26]
改善神经炎症[27]
促进神经发生[28]GLUT4 心肌[29]、骨骼肌[15]、
脂肪[30]囊泡转运到细胞膜上[23] 参与神经元葡萄糖代谢[31]
改善中枢胰岛素敏感性[32]成纤维生长因子21(Fibroblast Growth Factor 21, FGF 21) 肝脏[33]、脂肪[34]、
肌肉[35]以游离态通过血液循环转运[36] 改善神经炎症[37]
应对应激,调节代谢[38]
调节摄食行为[39]
促进中枢神经系统髓鞘再生[40]
促进神经元发育[41]IGF-1 肝脏[42]、骨骼肌[43] 与IGFBP结合形成复合物在血液中运输[44],与靶细胞膜上的特异性受体结合进行信号转导[45] 促进早期大脑功能及发育[46]
参与神经保护,塑造大脑结构[47]
促进神经可塑性,有助于学习与记忆[48]
改善脑血管功能[49]BDNF 脑[19]、骨骼肌[50] 以游离态经过血液循环转运[51]
由囊泡运输[52]减少神经元凋亡,缓解认知障碍[53]
恢复突触可塑性,改善神经发生,减少神经炎症[54]VEGF 成纤维细胞[55]、平滑肌细胞[56]、小胶质细胞[57] 以游离态在血液循环中转运[58]
与结合蛋白形成复合物在血液中转运[59]刺激神经发生[60]
改善神经炎症[61]
促进脑血管生成[62]FNDC5 肌肉[63]、肝脏[64]、
脂肪[65]在骨骼肌细胞中,FNDC5先剪切、修饰后生成鸢尾素,鸢尾素再分泌至血液中,经血液循环运送至靶向部位发挥作用[20] 提高突触可塑性[66]
促进神经营养因子表达[66]
改善神经炎症[66]乳酸 肌肉、肝脏、脑[67] 经过乳酸转运蛋白转运[68]
经血液循环运输促进胶质细胞与神经元之间的能量传递[69]
维持正常的突触功能[70]
改善神经炎症,缓解认知功能障碍[14]MicroRNA miR-382-3p
miR-186
miR-146a
miR-133a/b
miR-532-5p几乎所有器官和组织 囊泡运输、胞吐、脂质体运输 改善认知障碍[71]
改善神经元凋亡[72]
减少大脑炎症[73]
提高胰岛素敏感性[74]
改善认知功能[75] 下载: 导出CSV
下载: 导出CSV
-
[1] GBD 2019 Mental Disorders Collaborations. Global,regional,and national burden of 12 mental disorders in 204 countries and territories,1990-2019:A systematic analysis for the Global Burden of Disease Study 2019[J]. The Lancet Psychiatry,2022,9(2):137-150 doi: 10.1016/S2215-0366(21)00395-3
[2] FERNANDO A P,ROBBINS T W. Animal models of neuropsychiatric disorders[J]. Annual Review of Clinical Psychology,2011,7:39-61 doi: 10.1146/annurev-clinpsy-032210-104454
[3] SWENSON S,BLUM K,MCLAUGHLIN T,et al. The therapeutic potential of exercise for neuropsychiatric diseases:A review[J]. Journal of the Neurological Sciences,2020,412:116763 doi: 10.1016/j.jns.2020.116763
[4] HE W K,ZHANG S,QI Z T,et al. Unveiling the potential of estrogen:Exploring its role in neuropsychiatric disorders and exercise intervention[J]. Pharmacological Research,2024,204:107201 doi: 10.1016/j.phrs.2024.107201
[5] CHOW L S,GERSZTEN R E,TAYLOR J M,et al. Exerkines in health,resilience and disease[J]. Nature Reviews. Endocrinology,2022,18(5):273-289 doi: 10.1038/s41574-022-00641-2
[6] ISLAM M R,VALARIS S,YOUNG M F,et al. Exercise hormone irisin is a critical regulator of cognitive function[J]. Nature Metabolism,2021,3(8):1058-1070 doi: 10.1038/s42255-021-00438-z
[7] EL HAYEK L,KHALIFEH M,ZIBARA V,et al. Lactate mediates the effects of exercise on learning and memory through SIRT1-dependent activation of hippocampal brain-derived neurotrophic factor (BDNF)[J]. The Journal of Neuroscience,2019,39(13):2369-2382
[8] PEDERSEN B K,FISCHER C P. Beneficial health effects of exercise:The role of IL-6 as a myokine[J]. Trends in Pharmacological Sciences,2007,28(4):152-156 doi: 10.1016/j.tips.2007.02.002
[9] 饶志坚,张建红,赵杰修. 运动因子调控网络分析[J]. 中国运动医学杂志,2023,42(12):987-1000 doi: 10.3969/j.issn.1000-6710.2023.12.011 [10] 张若琳,漆正堂,刘微娜. 微生物-肠-脑轴视角下色氨酸代谢介导的运动抗抑郁机制研究进展[J]. 中国运动医学杂志,2023,42(3):227-235 doi: 10.3969/j.issn.1000-6710.2023.03.008 [11] PEDERSEN B K,STEENSBERG A,FISCHER C,et al. Searching for the exercise factor:Is IL-6 a candidate?[J]. Journal of Muscle Research and Cell Motility,2003,24(2-3):113-119
[12] PEDERSEN B K,FEBBRAIO M. Muscle-derived interleukin-6:A possible link between skeletal muscle,adipose tissue,liver,and brain[J]. Brain,Behavior,and Immunity,2005,19(5):371-376
[13] MOON H Y,BECKE A,BERRON D,et al. Running-induced systemic cathepsin B secretion is associated with memory function[J]. Cell Metabolism,2016,24(2):332-340 doi: 10.1016/j.cmet.2016.05.025
[14] HAN H,ZHAO Y W,DU J D,et al. Exercise improves cognitive dysfunction and neuroinflammation in mice through Histone H3 lactylation in microglia[J]. Immunity & Ageing,2023,20(1):63
[15] 王平,李佳欣,陈小龙,等. 转录因子EB在有氧运动改善高脂饮食诱导小鼠骨骼肌胰岛素抵抗中的作用[J]. 中国运动医学杂志,2024,43(3):193-204 doi: 10.3969/j.issn.1000-6710.2024.03.007 [16] 税晓平,李春莹,李顺昌,等. 有氧和抗阻运动干预2型糖尿病大鼠骨骼肌脑源性神经营养因子、核因子κB及炎症指标的表达[J]. 中国组织工程研究,2022,26(5):669-675 doi: 10.12307/2022.109 [17] PARK H,POO M M. Neurotrophin regulation of neural circuit development and function[J]. Nature Reviews. Neuroscience,2013,14(1):7-23 doi: 10.1038/nrn3379
[18] 张康,张业廷,付燕. 跑台运动对帕金森病模型小鼠不同脑区DAT、BDNF和TrkB表达的影响[J]. 中国运动医学杂志,2022,41(9):704-713 doi: 10.3969/j.issn.1000-6710.2022.09.006 [19] 陆小香,张蕴琨,江年. 力竭运动后大鼠海马CA1区自由基、下丘脑GABA及HPA轴的动态变化[J]. 中国运动医学杂志,2007,26(5):563-567 doi: 10.3969/j.issn.1000-6710.2007.05.010 [20] LOURENCO M V,FROZZA R L,DE FREITAS G B,et al. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer's models[J]. Nature Medicine,2019,25(1):165-175 doi: 10.1038/s41591-018-0275-4
[21] WANG D C,LIN H T,LEE Y J,et al. Recovery of BDNF and CB1R in the prefrontal cortex underlying improvement of working memory in prenatal DEHP-exposed male rats after aerobic exercise[J]. International Journal of Molecular Sciences,2020,21(11):3867 doi: 10.3390/ijms21113867
[22] 刘文彬,刘微娜,漆正堂. 神经营养因子介导运动的抗抑郁作用[J]. 体育科学,2018,38(10):54-66 [23] 崔菊,庞静,宫环,等. GLUT4和脂联素在3T3-L1脂肪细胞中共享囊泡转运机制[J]. 医学研究杂志,2017,46(3):18-21 doi: 10.11969/j.issn.1673-548X.2017.03.006 [24] 王少堃,王世强,王一杰,等. 骨骼肌介导的运动神经保护效应:作用途径和分子机制[J]. 中国体育科技,2023,59(4):58-66 [25] ZHANG S H,WU X,WANG J,et al. Adiponectin/AdiopR1 signaling prevents mitochondrial dysfunction and oxidative injury after traumatic brain injury in a SIRT3 dependent manner[J]. Redox Biology,2022,54:102390 doi: 10.1016/j.redox.2022.102390
[26] 田清华,刘霞,邓鹏辉,等. 有氧运动对糖尿病大鼠学习记忆功能、海马突触可塑性及脂联素信号通路的影响[J]. 中华老年医学杂志,2024,43(3):348-353 doi: 10.3760/cma.j.issn.0254-9026.2024.03.014 [27] LI W F,ALI T,ZHENG C Y,et al. Anti-depressive-like behaviors of APN KO mice involve Trkb/BDNF signaling related neuroinflammatory changes[J]. Molecular Psychiatry,2022,27(2):1047-1058 doi: 10.1038/s41380-021-01327-3
[28] LEE T H,CHRISTIE B R,VAN PRAAG H,et al. AdipoRon treatment induces a dose-dependent response in adult hippocampal neurogenesis[J]. International Journal of Molecular Sciences,2021,22(4):2068 doi: 10.3390/ijms22042068
[29] 李欣,余福林,高延. 跑台运动调节线粒体ATP释放改善高脂饮食大鼠的心肌损伤[J]. 心脏杂志,2024,36(3):256-261 doi: 10.12125/j.chj.202304119 [30] 吴彪,范冬霞,张佳,等. PM2.5和热/冷暴露对小鼠骨骼肌和白色脂肪组织AKT/GLUT4通路的影响[J]. 环境与职业医学. 2024,41(4):356-361 [31] PENG W X,TAN C H,MO L J,et al. Glucose transporter 3 in neuronal glucose metabolism:Health and diseases[J]. Metabolism,2021,123:154869 doi: 10.1016/j.metabol.2021.154869
[32] YONAMINE C Y,MICHALANI M L E,MOREIRA R J,et al. Glucose transport and utilization in the hippocampus:From neurophysiology to diabetes-related development of dementia[J]. International Journal of Molecular Sciences,2023,24(22):16480 doi: 10.3390/ijms242216480
[33] 张媛,盛蕾,刘小玮,等. 不同运动方式对肥胖大鼠肝脏脂质沉积及FGF21分泌的影响[J]. 中国应用生理学杂志,2022,38(1):47-52 [34] GENG L L,LIAO B Y,JIN L G,et al. Exercise alleviates obesity-induced metabolic dysfunction via enhancing FGF21 sensitivity in adipose tissues[J]. Cell Reports,2019,26(10):2738-2752
[35] GAO Y,ZHANG W,ZENG L Q,et al. Exercise and dietary intervention ameliorate high-fat diet-induced NAFLD and liver aging by inducing lipophagy[J]. Redox Biology,2020,36:101635 doi: 10.1016/j.redox.2020.101635
[36] 李良,徐建方,冯连世,等. 有氧运动和抗阻运动训练对肥胖大鼠肝脏FGF21信号通路的影响[J]. 中国运动医学杂志,2018,37(10):847-856 doi: 10.3969/j.issn.1000-6710.2018.10.007 [37] WANG D X,LIU F,ZHU L Y,et al. FGF21 alleviates neuroinflammation following ischemic stroke by modulating the temporal and spatial dynamics of microglia/macrophages[J]. Journal of Neuroinflammation,2020,17(1):257 doi: 10.1186/s12974-020-01921-2
[38] GENG L L,LAM K S L,XU A M. The therapeutic potential of FGF21 in metabolic diseases:From bench to clinic[J]. Nature Reviews. Endocrinology,2020,16(11):654-667 doi: 10.1038/s41574-020-0386-0
[39] FLIPPO K H,TRAMMELL S A J,GILLUM M P,et al. FGF21 suppresses alcohol consumption through an amygdalo-striatal circuit[J]. Cell Metabolism,2022,34(2):317-328
[40] KURODA M,MURAMATSU R,MAEDERA N,et al. Peripherally derived FGF21 promotes remyelination in the central nervous system[J]. The Journal of Clinical Investigation,2017,127(9):3496-3509 doi: 10.1172/JCI94337
[41] 李晶晶,孙晓彤,柳婷婷,等. 成纤维细胞生长因子21与妊娠期代谢性疾病的研究进展[J]. 国际妇产科学杂志,2023,50(4):400-404 doi: 10.12280/gjfckx.20230156 [42] 黄嵩,王巍,牛燕媚,等. 有氧运动干预父系C57BL/6小鼠对雄性子代IGF-1表达及其甲基化的影响[J]. 中国运动医学杂志,2020,39(4):288-294 doi: 10.3969/j.issn.1000-6710.2020.04.006 [43] 刘延莹,杨海平,冯庆鲲,等. 弱激光预照射联合有氧运动对增龄大鼠骨骼肌SIRTs/PGC-1α轴及IGF-1蛋白表达的影响[J]. 中国运动医学杂志,2019,38(7):577-584 doi: 10.3969/j.issn.1000-6710.2019.07.006 [44] HUANG R R,SHI J Y,WEI R H,et al. Challenges of insulin-like growth factor-1 testing[J]. Critical Reviews in Clinical Laboratory Sciences,2024,61(5):388-403 doi: 10.1080/10408363.2024.2306804
[45] 程慧芳,李会会,王庆志,等. 迷走神经刺激对老龄小鼠术后认知功能障碍的影响及海马IGF-1信号通路在其中的作用[J]. 中华麻醉学杂志,2022(9):1048-1053 doi: 10.3760/cma.j.cn131073.20220617.00906 [46] RUSIN D,VAHL BECIROVIC L,LYSZCZARZ G,et al. Microglia-derived insulin-like growth factor 1 is critical for neurodevelopment[J]. Cells,2024,13(2):184 doi: 10.3390/cells13020184
[47] NUÑEZ A,ZEGARRA-VALDIVIA J,FERNANDEZ DE SEVILLA D,et al. The neurobiology of insulin-like growth factor I:From neuroprotection to modulation of brain states[J]. Molecular Psychiatry,2023,28(8):3220-3230 doi: 10.1038/s41380-023-02136-6
[48] MAGLIO L E,NORIEGA-PRIETO J A,MAROTO I B,et al. IGF-1 facilitates extinction of conditioned fear[J]. eLife,2021,10:e67267 doi: 10.7554/eLife.67267
[49] ZHANG J Y,LIU M Q,HUANG M H,et al. Ginsenoside F1 promotes angiogenesis by activating the IGF-1/IGF1R pathway[J]. Pharmacological Research,2019,144:292-305 doi: 10.1016/j.phrs.2019.04.021
[50] CAVALERI D,MORETTI F,BARTOCCETTI A,et al. The role of BDNF in major depressive disorder,related clinical features,and antidepressant treatment:Insight from meta-analyses[J]. Neuroscience & Biobehavioral Reviews,2023,149:105159
[51] REED J L,TERADA T,COTIE L M,et al. The effects of high-intensity interval training,Nordic walking and moderate-to-vigorous intensity continuous training on functional capacity,depression and quality of life in patients with coronary artery disease enrolled in cardiac rehabilitation:A randomized controlled trial (CRX study)[J]. Progress in Cardiovascular Diseases,2022,70:73-83 doi: 10.1016/j.pcad.2021.07.002
[52] DELGADO-PERAZA F,NOGUERAS-ORTIZ C,SIMONSEN A H,et al. Neuron-derived extracellular vesicles in blood reveal effects of exercise in Alzheimer's disease[J]. Alzheimer's Research & Therapy,2023,15(1):156
[53] WANG J,NIU Y L,TAO H Y,et al. Knockdown of lncRNA TUG1 inhibits hippocampal neuronal apoptosis and participates in aerobic exercise-alleviated vascular cognitive impairment[J]. Biological Research,2020,53(1):53 doi: 10.1186/s40659-020-00320-4
[54] ZHOU X,DENG X H,LIU M F,et al. Intranasal delivery of BDNF-loaded small extracellular vesicles for cerebral ischemia therapy[J]. Journal of Controlled Release,2023,357:1-19 doi: 10.1016/j.jconrel.2023.03.033
[55] CADAMURO M,BRIVIO S,MERTENS J,et al. Platelet-derived growth factor-D enables liver myofibroblasts to promote tumor lymphangiogenesis in cholangiocarcinoma[J]. Journal of Hepatology,2019,70(4):700-709 doi: 10.1016/j.jhep.2018.12.004
[56] GIELEN S,SANDRI M,ERBS S,et al. Exercise-induced modulation of endothelial nitric oxide production[J]. Current Pharmaceutical Biotechnology,2011,12(9):1375-1384 doi: 10.2174/138920111798281063
[57] CHOI B R,JOHNSON K R,MARIC D,et al. Monocyte-derived IL-6 programs microglia to rebuild damaged brain vasculature[J]. Nature Immunology,2023,24(7):1110-1123 doi: 10.1038/s41590-023-01521-1
[58] MORLAND C,ANDERSSON K A,HAUGEN Ø P,et al. Exercise induces cerebral VEGF and angiogenesis via the lactate receptor HCAR1[J]. Nature Communications,2017,8:15557 doi: 10.1038/ncomms15557
[59] GOEL S,DUDA D,XU L,et al. Normalization of the vasculature for treatment of cancer and other diseases[J]. Physiological Reviews,2011,91(3):1071-1121 doi: 10.1152/physrev.00038.2010
[60] DU PREEZ A,ONORATO D,EIBEN I,et al. Chronic stress followed by social isolation promotes depressive-like behaviour,alters microglial and astrocyte biology and reduces hippocampal neurogenesis in male mice[J]. Brain,Behavior,and Immunity,2021,91:24-47
[61] BOISSERAND L S B,GERALDO L H,BOUCHART J,et al. VEGF-C prophylaxis favors lymphatic drainage and modulates neuroinflammation in a stroke model[J]. The Journal of Experimental Medicine,2024,221(4):e20221983 doi: 10.1084/jem.20221983
[62] ZONG X M,LI Y Y,LIU C,et al. Theta-burst transcranial magnetic stimulation promotes stroke recovery by vascular protection and neovascularization[J]. Theranostics,2020,10(26):12090-12110 doi: 10.7150/thno.51573
[63] KIM H,WRANN C D,JEDRYCHOWSKI M,et al. Irisin mediates effects on bone and fat via αV integrin receptors[J]. Cell,2018,175(7):1756-1768
[64] LI D J,SUN S J,FU J T,et al. NAD+-boosting therapy alleviates nonalcoholic fatty liver disease via stimulating a novel exerkine Fndc5/irisin[J]. Theranostics,2021,11(9):4381-4402 doi: 10.7150/thno.53652
[65] HE X Y,HUA Y,LI Q,et al. FNDC5/irisin facilitates muscle−adipose−bone connectivity through ubiquitination-dependent activation of runt-related transcriptional factors RUNX1/2[J]. Journal of Biological Chemistry,2022,298(3):101679 doi: 10.1016/j.jbc.2022.101679
[66] 章森,邹勇,漆正堂,等. 鸢尾素介导运动干预神经精神疾病的潜在机制[J]. 上海体育学院学报,2023,47(4):39-50 [67] MAGISTRETTI P J,ALLAMAN I. Lactate in the brain:From metabolic end-product to signalling molecule[J]. Nature Reviews. Neuroscience,2018,19(4):235-249 doi: 10.1038/nrn.2018.19
[68] WANG N,JIANG X,ZHANG S,et al. Structural basis of human monocarboxylate transporter 1 inhibition by anti-cancer drug candidates[J]. Cell,2021,184(2):370-383
[69] LIU L,MACKENZIE K R,PUTLURI N,et al. The glia-neuron lactate shuttle and elevated ROS promote lipid synthesis in neurons and lipid droplet accumulation in glia via APOE/D[J]. Cell Metabolism,2017,26(5):719-737
[70] MONSORNO K,GINGGEN K,IVANOV A,et al. Loss of microglial MCT4 leads to defective synaptic pruning and anxiety-like behavior in mice[J]. Nature Communications,2023,14(1):5749 doi: 10.1038/s41467-023-41502-4
[71] WANG M Z,XIE K L,ZHAO S N,et al. Aerobic exercise improves cognitive impairment in mice with type 2 diabetes by regulating the MALAT1/miR-382-3p/BDNF signaling pathway in serum-exosomes[J]. Molecular Medicine,2023,29(1):130 doi: 10.1186/s10020-023-00727-1
[72] NIU Y L,WAN C X,ZHANG J,et al. Aerobic exercise improves VCI through circRIMS2/miR-186/BDNF-mediated neuronal apoptosis[J]. Molecular Medicine,2021,27(1):4 doi: 10.1186/s10020-020-00258-z
[73] ZHANG Z F,ZOU X X,ZHANG R,et al. Human umbilical cord mesenchymal stem cell-derived exosomal miR-146a-5p reduces microglial-mediated neuroinflammation via suppression of the IRAK1/TRAF6 signaling pathway after ischemic stroke[J]. Aging,2021,13(2):3060-3079 doi: 10.18632/aging.202466
[74] CASTAÑO C,MIRASIERRA M,VALLEJO M,et al. Delivery of muscle-derived exosomal miRNAs induced by HIIT improves insulin sensitivity through down-regulation of hepatic FoxO1 in mice[J]. Proceedings of the National Academy of Sciences of the United States of America,2020,117(48):30335-30343
[75] LIANG X Y,FA W X,WANG N,et al. Exosomal miR-532-5p induced by long-term exercise rescues blood–brain barrier function in 5XFAD mice via downregulation of EPHA4[J]. Aging Cell,2023,22(1):e13748 doi: 10.1111/acel.13748
[76] KIM H J,SONG B K,SO B,et al. Increase of circulating BDNF levels and its relation to improvement of physical fitness following 12 weeks of combined exercise in chronic patients with schizophrenia:A pilot study[J]. Psychiatry Research,2014,220(3):792-796 doi: 10.1016/j.psychres.2014.09.020
[77] ZHANG J Q,RONG P J,ZHANG L J,et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis[J]. Science Advances,2021,7(12):eabb9888 doi: 10.1126/sciadv.abb9888
[78] 王璐,闫冬,夏瑞洁,等. 运动预处理通过外泌体介导miR-146a对大鼠缺血性脑卒中炎症反应的影响[J]. 中国运动医学杂志,2023,42(11):889-898 doi: 10.3969/j.issn.1000-6710.2023.11.007 [79] DEL GIUDICE M,GANGESTAD S W. Rethinking IL-6 and CRP:Why they are more than inflammatory biomarkers,and why it matters[J]. Brain,Behavior,and Immunity,2018,70:61-75
[80] XU H,WANG J S,ZHOU Y J,et al. BDNF affects the mediating effect of negative symptoms on the relationship between age of onset and cognition in patients with chronic schizophrenia[J]. Psychoneuroendocrinology,2021,125:105121 doi: 10.1016/j.psyneuen.2020.105121
[81] 王丽. 天麻素片联合盐酸帕罗西汀对老年脑卒中后焦虑抑郁患者血清BDNF、IGF-1浓度的影响[J]. 中国老年学杂志,2017,37(21):5403-5405 doi: 10.3969/j.issn.1005-9202.2017.21.083 [82] CHEN Y S,CHEN X F,LUO Z W,et al. Exercise-induced reduction of IGF1R sumoylation attenuates neuroinflammation in APP/PS1 transgenic mice[J]. Journal of Advanced Research,2024,3:25
[83] ESTEVES M,ABREU R,FERNANDES H,et al. MicroRNA-124-3p-enriched small extracellular vesicles as a therapeutic approach for Parkinson's disease[J]. Molecular Therapy,2022,30(10):3176-3192 doi: 10.1016/j.ymthe.2022.06.003
[84] CAO Z,MIN J H,TAN Q L,et al. Circulating insulin-like growth factor-1 and brain health:Evidence from 369,711 participants in the UK Biobank[J]. Alzheimer's Research & Therapy,2023,15(1):140
[85] PINTO-SANCHEZ M I,HALL G B,GHAJAR K,et al. Probiotic bifidobacterium longum NCC3001 reduces depression scores and alters brain activity:A pilot study in patients with irritable bowel syndrome[J]. Gastroenterology,2017,153(2):448-459
[86] KHAN A S,PETERSON K A,VITTAY O I,et al. Deuterium metabolic imaging of Alzheimer disease at 3-T magnetic field strength:A pilot case-control study[J]. Radiology,2024,312(1):e232407 doi: 10.1148/radiol.232407
[87] TIJMS B M,VROMEN E M,MJAAVATTEN O,et al. Cerebrospinal fluid proteomics in patients with Alzheimer's disease reveals five molecular subtypes with distinct genetic risk profiles[J]. Nature Aging,2024,4(1):33-47 doi: 10.1038/s43587-023-00550-7
[88] PHD M D,MSC P P,MSC C Z,et al. Irisin levels in cerebrospinal fluid correlate with biomarkers and clinical dementia scores in Alzheimer disease[J]. Annals of Neurology,2024,96(1):61-73 doi: 10.1002/ana.26946
[89] SZUHANY K L,OTTO M W. Assessing BDNF as a mediator of the effects of exercise on depression[J]. Journal of Psychiatric Research,2020,123:114-118 doi: 10.1016/j.jpsychires.2020.02.003
[90] XIA J,XUE X L,LIU W B,et al. The role of Fgf9 in the antidepressant effects of exercise and fluoxetine in chronic unpredictable mild stress mice[J]. Psychosomatic Medicine,2021,83(7):795-804 doi: 10.1097/PSY.0000000000000953
[91] LIU R,WANG X D,AIHARA K,et al. Early diagnosis of complex diseases by molecular biomarkers,network biomarkers,and dynamical network biomarkers[J]. Medicinal Research Reviews,2014,34(3):455-478 doi: 10.1002/med.21293
[92] CONTREPOIS K,WU S,MONEGHETTI K J,et al. Molecular choreography of acute exercise[J]. Cell,2020,181(5):1112-1130
[93] VETR N G,GAY N R,GROUP M S,et al. The impact of exercise on gene regulation in association with complex trait genetics[J]. Nature Communications,2024,15(1):3346 doi: 10.1038/s41467-024-45966-w
[94] MANY G M,SANFORD J A,SAGENDORF T J,et al. Sexual dimorphism and the multi-omic response to exercise training in rat subcutaneous white adipose tissue[J]. Nature Metabolism,2024,6:963-979 doi: 10.1038/s42255-023-00959-9
[95] WANG Z,EMMERICH A,PILLON N J,et al. Genome-wide association analyses of physical activity and sedentary behavior provide insights into underlying mechanisms and roles in disease prevention[J]. Nature Genetics,2022,54(9):1332-1344 doi: 10.1038/s41588-022-01165-1
[96] YAN L,WEI J,YANG F Z,et al. Physical exercise prevented stress-induced anxiety via improving brain RNA methylation (adv. sci. 24/2022)[J]. Advanced Science,2022,9(24):2270148 doi: 10.1002/advs.202270148
[97] DE MIGUEL Z,KHOURY N,BETLEY M J,et al. Exercise plasma boosts memory and dampens brain inflammation via clusterin[J]. Nature,2021,600(7889):494-499 doi: 10.1038/s41586-021-04183-x
[98] LIU W N,HUANG Z C,ZHANG Y,et al. ASMT determines gut microbiota and increases neurobehavioral adaptability to exercise in female mice[J]. Communications Biology,2023,6(1):1126 doi: 10.1038/s42003-023-05520-8
[99] 章森,刘文彬,夏杰,等. 运动改善ASMT基因敲除小鼠抑郁行为的海马蛋白质组学机制[J]. 上海体育大学学报,2024,48(3):36-48 [100] KALAY Z. Fundamental and functional aspects of mesoscopic architectures with examples in physics,cell biology,and chemistry[J]. Critical Reviews in Biochemistry and Molecular Biology,2011,46(4):310-326 doi: 10.3109/10409238.2011.582081
[101] LI X N,YU B,SUN Q T,et al. Generation of a whole-brain atlas for the cholinergic system and mesoscopic projectome analysis of basal forebrain cholinergic neurons[J]. Proceedings of the National Academy of Sciences of the United States of America,2018,115(2):415-420
[102] WALTER T J,SUTER R K,AYAD N G. An overview of human single-cell RNA sequencing studies in neurobiological disease[J]. Neurobiology of Disease,2023,184:106201 doi: 10.1016/j.nbd.2023.106201
[103] CATHOMAS F,LIN H Y,CHAN K L,et al. Circulating myeloid-derived MMP8 in stress susceptibility and depression[J]. Nature,2024,626(8001):1108-1115 doi: 10.1038/s41586-023-07015-2
[104] NAGY C,MAITRA M,TANTI A,et al. Single-nucleus transcriptomics of the prefrontal cortex in major depressive disorder implicates oligodendrocyte precursor cells and excitatory neurons[J]. Nature Neuroscience,2020,23(6):771-781 doi: 10.1038/s41593-020-0621-y
[105] SUN S H,MA S,CAI Y S,et al. A single-cell transcriptomic atlas of exercise-induced anti-inflammatory and geroprotective effects across the body[J]. The Innovation,2023,4(1):100380 doi: 10.1016/j.xinn.2023.100380
[106] JIN K,YAO Z Z,VAN VELTHOVEN C T J,et al. Cell-type specific molecular signatures of aging revealed in a brain-wide transcriptomic cell-type atlas[J]. bioRxiv,2023:2023.07. 26.550355
[107] LANGLIEB J,SACHDEV N S,BALDERRAMA K S,et al. The cell type composition of the adult mouse brain revealed by single cell and spatial genomics[J]. bioRxiv,2023:2023.03. 06.531307
[108] KATHE C,SKINNIDER M A,HUTSON T H,et al. The neurons that restore walking after paralysis[J]. Nature,2022,611(7936):540-547 doi: 10.1038/s41586-022-05385-7
[109] HUH J Y,MOUGIOS V,SKRAPARLIS A,et al. Irisin in response to acute and chronic whole-body vibration exercise in humans[J]. Metabolism,2014,63(7):918-921 doi: 10.1016/j.metabol.2014.04.001
[110] TSUCHIYA Y,ANDO D,TAKAMATSU K,et al. Resistance exercise induces a greater irisin response than endurance exercise[J]. Metabolism,2015,64(9):1042-1050 doi: 10.1016/j.metabol.2015.05.010
[111] TSAI C L,PAN C Y,TSENG Y T,et al. Acute effects of high-intensity interval training and moderate-intensity continuous exercise on BDNF and irisin levels and neurocognitive performance in late middle-aged and older adults[J]. Behavioural Brain Research,2021,413:113472 doi: 10.1016/j.bbr.2021.113472
[112] REYCRAFT J T,ISLAM H,TOWNSEND L K,et al. Exercise intensity and recovery on circulating brain-derived neurotrophic factor[J]. Medicine and Science in Sports and Exercise,2020,52(5):1210-1217 doi: 10.1249/MSS.0000000000002242
[113] PAHLAVANI H A. Exercise therapy to prevent and treat Alzheimer's disease[J]. Frontiers in Aging Neuroscience,2023,15:1243869 doi: 10.3389/fnagi.2023.1243869
[114] DORA K,TSUKAMOTO H,SUGA T,et al. Essential amino acid supplements ingestion has a positive effect on executive function after moderate-intensity aerobic exercise[J]. Scientific Reports,2023,13(1):22644 doi: 10.1038/s41598-023-49781-z
[115] SUZUKI G,TOKUNO S,NIBUYA M,et al. Decreased plasma brain-derived neurotrophic factor and vascular endothelial growth factor concentrations during military training[J]. PLoS One,2014,9(2):e89455 doi: 10.1371/journal.pone.0089455
[116] MORRIS T P,BURZYNSKA A,VOSS M,et al. Brain structure and function predict adherence to an exercise intervention in older adults[J]. Medicine and Science in Sports and Exercise,2022,54(9):1483-1492 doi: 10.1249/MSS.0000000000002949
[117] MATA J,THOMPSON R J,GOTLIB I H. BDNF genotype moderates the relation between physical activity and depressive symptoms[J]. Health Psychology,2010,29(2):130-133 doi: 10.1037/a0017261
[118] RAHMAN M S,MILLISCHER V,ZEEBARI Z,et al. BDNF Val66Met and childhood adversity on response to physical exercise and Internet-based cognitive behavioural therapy in depressed Swedish adults[J]. Journal of Psychiatric Research,2017,93:50-58 doi: 10.1016/j.jpsychires.2017.05.007
[119] PITTS B L,WHEALIN J M,HARPAZ-ROTEM I,et al. BDNF Val66Met polymorphism and posttraumatic stress symptoms in U. S. military veterans:Protective effect of physical exercise[J]. Psychoneuroendocrinology,2019,100:198-202 doi: 10.1016/j.psyneuen.2018.10.011
[120] BRYANT R A,DAWSON K S,AZEVEDO S,et al. A pilot study of the role of the BDNF Val66Met polymorphism in response to exercise-augmented exposure therapy for posttraumatic stress disorder[J]. Psychoneuroendocrinology,2024,167:107106 doi: 10.1016/j.psyneuen.2024.107106
[121] PITTS B L,WEN V,WHEALIN J M,et al. Depression and cognitive dysfunction in older U. S. military veterans:Moderating effects of BDNF Val66Met polymorphism and physical exercise[J]. The American Journal of Geriatric Psychiatry,2020,28(9):959-967 doi: 10.1016/j.jagp.2020.02.001
[122] REICHEL T,HACKER S,PALMOWSKI J,et al. Neurophysiological markers for monitoring exercise and recovery cycles in endurance sports[J]. Journal of Sports Science & Medicine,2022,21(3):446-457
[123] REICHEL T,BOßLAU T K,PALMOWSKI J,et al. Reliability and suitability of physiological exercise response and recovery markers[J]. Scientific Reports,2020,10(1):11924 doi: 10.1038/s41598-020-69280-9
[124] LEE S,NOURAEIN S,KWON J J,et al. Engineered serum markers for non-invasive monitoring of gene expression in the brain[J]. Nature Biotechnology,2024,42(11):1717-1725 doi: 10.1038/s41587-023-02087-x
[125] KONING E,VORSTMAN J,MCINTYRE R S,et al. Characterizing eating behavioral phenotypes in mood disorders:A narrative review[J]. Psychological Medicine,2022,52(14):2885-2898 doi: 10.1017/S0033291722002446
[126] WANG J,TANG J,LIANG X,et al. Hippocampal PGC-1α-mediated positive effects on parvalbumin interneurons are required for the antidepressant effects of running exercise[J]. Translational Psychiatry,2021,11(1):222 doi: 10.1038/s41398-021-01339-1
[127] LIU W N,LIU J T,HUANG Z C,et al. Possible role of GLP-1 in antidepressant effects of metformin and exercise in CUMS mice[J]. Journal of Affective Disorders,2019,246:486-497 doi: 10.1016/j.jad.2018.12.112
[128] LIU W N,LIU J T,XIA J,et al. Leptin receptor knockout-induced depression-like behaviors and attenuated antidepressant effects of exercise are associated with STAT3/SOCS3 signaling[J]. Brain,Behavior,and Immunity,2017,61:297-305
[129] 黄卓淳,漆正堂,刘微娜. 运动与褪黑素抗抑郁的作用、机制以及联合策略研究进展[J]. 中国体育科技,2020,56(2):3-14 [130] BASTIOLI G,ARNOLD J C,MANCINI M,et al. Voluntary exercise boosts striatal dopamine release:Evidence for the necessary and sufficient role of BDNF[J]. The Journal of Neuroscience,2022,42(23):4725-4736 doi: 10.1523/JNEUROSCI.2273-21.2022
[131] HOROWITZ A M,FAN X L,BIERI G,et al. Blood factors transfer beneficial effects of exercise on neurogenesis and cognition to the aged brain[J]. Science,2020,369(6500):167-173 doi: 10.1126/science.aaw2622
[132] CHEN K,ZHENG Y H,WEI J,et al. Exercise training improves motor skill learning via selective activation of mTOR[J]. Science Advances,2019,5(7):eaaw1888 doi: 10.1126/sciadv.aaw1888
[133] ZHANG W J,OU H N,ZHANG B R,et al. Treadmill exercise relieves chronic restraint stress-induced cognitive impairments in mice via activating protein phosphatase 2A[J]. Neuroscience Bulletin,2021,37(10):1487-1492 doi: 10.1007/s12264-021-00766-w
[134] 陈平,李刚强,周文辉. 谷氨酸受体在帕金森病中的作用及其介导的帕金森病运动防治研究进展[J]. 体育科学,2022,42(9):72-81 [135] 陈巍,魏翔,刘晓莉,等. 运动对PD模型大鼠皮层-纹状体Glu能神经传递的影响[J]. 北京体育大学学报,2015,38(2):61-66 [136] 时凯旋,刘晓莉,乔德才. 运动通过调节皮层-纹状体通路功能连接可塑性改善PD模型大鼠行为[J]. 体育科学,2020,40(6):49-58 [137] CHENG T,HUANG X D,HU X F,et al. Physical exercise rescues cocaine-evoked synaptic deficits in motor cortex[J]. Molecular Psychiatry,2021,26(11):6187-6197 doi: 10.1038/s41380-021-01336-2
[138] HE Y,MADEO G,LIANG Y,et al. A red nucleus-VTA glutamate pathway underlies exercise reward and the therapeutic effect of exercise on cocaine use[J]. Science Advances,2022,8(35):eabo1440 doi: 10.1126/sciadv.abo1440
[139] JUNG S,KIM Y,KIM M,et al. Exercise pills for drug addiction:Forced moderate endurance exercise inhibits methamphetamine-induced hyperactivity through the striatal glutamatergic signaling pathway in male sprague dawley rats[J]. International Journal of Molecular Sciences,2021,22(15):8203 doi: 10.3390/ijms22158203
[140] DONG Y G,GAN Y X,FU Y M,et al. Treadmill exercise training inhibits morphine CPP by reversing morphine effects on GABA neurotransmission in D2-MSNs of the accumbens-pallidal pathway in male mice[J]. Neuropsychopharmacology,2024,49(11):1700-1710 doi: 10.1038/s41386-024-01869-4
[141] LIANG X,LI R,WONG S H S,et al. The effects of exercise interventions on executive functions in children and adolescents with autism spectrum disorder:A systematic review and meta-analysis[J]. Sports Medicine,2022,52(1):75-88 doi: 10.1007/s40279-021-01545-3
[142] 朱笑彤,毕小羽,朱飞龙,等. ADHD学龄儿童动作协调能力与其核心症状的关系:基于执行功能的多重中介效应分析[J]. 体育科学,2023,43(7):65-73 [143] 杨孟超,金鹏,王德新,等. 运动改善注意缺陷多动障碍儿童执行功能和注意力的研究进展[J]. 体育科学,2022,42(5):77-87 [144] 苏余,张韧仁,何毅. 中等强度运动积极影响注意缺陷多动障碍儿童的问题行为及心率变异性研究[J]. 中国体育科技,2023,59(5):43-51 [145] ZHAO P T,CHEN K,ZHU G H,et al. Effects of aquatic exercise intervention on executive function and brain-derived neurotrophic factor of children with autism spectrum disorder[J]. Research in Developmental Disabilities,2024,150:104759 doi: 10.1016/j.ridd.2024.104759
[146] KING C,ROGERS L G,JANSEN J,et al. Adolescent treadmill exercise enhances hippocampal brain-derived neurotrophic factor (BDNF) expression and improves cognition in autism-modeled rats[J]. Physiology & Behavior,2024,284:114638
[147] SIVAYOKAN B,KING C,MALI I,et al. Aerobic exercise improves cognitive flexibility and modulates regional volume changes in a rat model of autism[J]. Behavioural Brain Research,2024,471:115136 doi: 10.1016/j.bbr.2024.115136
[148] RAHIMI R,AKHAVAN M M,KAMYAB K,et al. Maternal voluntary exercise ameliorates learning deficit in rat pups exposed,in utero,to valproic acid; role of BDNF and VEGF and their receptors[J]. Neuropeptides,2018,71:43-53 doi: 10.1016/j.npep.2018.06.006
[149] CHOI J W,HAN D H,KANG K D,et al. Aerobic exercise and attention deficit hyperactivity disorder:Brain research[J]. Medicine and Science in Sports and Exercise,2015,47(1):33-39 doi: 10.1249/MSS.0000000000000373
[150] TOSTA A,FONSECA A S,MESSEDER D,et al. Effects of gestational exercise on nociception,BDNF,and irisin levels in an animal model of ADHD[J]. Neuroscience,2024,543:37-48 doi: 10.1016/j.neuroscience.2024.02.008
[151] JEONG H I,JI E S,KIM S H,et al. Treadmill exercise improves spatial learning ability by enhancing brain-derived neurotrophic factor expression in the attention-deficit/hyperactivity disorder rats[J]. Journal of Exercise Rehabilitation,2014,10(3):162-167 doi: 10.12965/jer.140111
[152] WAKEFIELD L,AGARWAL S,TANNER K. Preclinical models for drug discovery for metastatic disease[J]. Cell,2023,186(8):1792-1813 doi: 10.1016/j.cell.2023.02.026
[153] COLEY N,GIULIOLI C,AISEN P S,et al. Randomised controlled trials for the prevention of cognitive decline or dementia:A systematic review[J]. Ageing Research Reviews,2022,82:101777 doi: 10.1016/j.arr.2022.101777
[154] SALMERON B J. Heterogeneity exists in healthy populations as well as in neuropsychiatric disorders[J]. Biological Psychiatry. Cognitive Neuroscience and Neuroimaging,2021,6(5):501-502 doi: 10.1016/j.bpsc.2021.03.002
[155] SCHELTENS P,DE STROOPER B,KIVIPELTO M,et al. Alzheimer's disease[J]. Lancet,2021,397(10284):1577-1590 doi: 10.1016/S0140-6736(20)32205-4
[156] LU B,CHEN X,XAVIER CASTELLANOS F,et al. The power of many brains:Catalyzing neuropsychiatric discovery through open neuroimaging data and large-scale collaboration[J]. Science Bulletin,2024,69(10):1536-1555 doi: 10.1016/j.scib.2024.03.006
[157] MULAY A R,HWANG J,KIM D H. Microphysiological blood-brain barrier systems for disease modeling and drug development[J]. Advanced Healthcare Materials,2024,13(21):2303180 doi: 10.1002/adhm.202303180
[158] NANCE E,PUN S H,SAIGAL R,et al. Drug delivery to the central nervous system[J]. Nature Reviews. Materials,2022,7(4):314-331
[159] JUTHANI R,MADAJEWSKI B,YOO B,et al. Ultrasmall core-shell silica nanoparticles for precision drug delivery in a high-grade malignant brain tumor model[J]. Clinical Cancer Research,2020,26(1):147-158 doi: 10.1158/1078-0432.CCR-19-1834
[160] KAPLAN A L,CONFAIR D N,KIM K,et al. Bespoke library docking for 5-HT2A receptor agonists with antidepressant activity[J]. Nature,2022,610(7932):582-591 doi: 10.1038/s41586-022-05258-z
[161] BRACHA S,JOHNSON H J,PRANCKEVICIUS N A,et al. Engineering Toxoplasma gondii secretion systems for intracellular delivery of multiple large therapeutic proteins to neurons[J]. Nature Microbiology,2024,9(8):2051-2072 doi: 10.1038/s41564-024-01750-6
[162] FAN W W,EVANS R M. Exercise mimetics:Impact on health and performance[J]. Cell Metabolism,2017,25(2):242-247 doi: 10.1016/j.cmet.2016.10.022
[163] WRANN C D,WHITE J P,SALOGIANNNIS J,et al. Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway[J]. Cell Metabolism,2013,18(5):649-659 doi: 10.1016/j.cmet.2013.09.008
[164] KOBILO T,YUAN C Y,VAN PRAAG H. Endurance factors improve hippocampal neurogenesis and spatial memory in mice[J]. Learning & Memory,2011,18(2):103-107
[165] CHAMBERLAIN S,GABRIEL H,STRITTMATTER W,et al. An exploratory phase Ⅱa study of the PPAR delta/gamma agonist T3D-959 assessing metabolic and cognitive function in subjects with mild to moderate Alzheimer's disease[J]. Journal of Alzheimer's Disease,2020,73(3):1085-1103 doi: 10.3233/JAD-190864


















计量
- 文章访问数: 1
- HTML全文浏览量: 0
- PDF下载量: 2